КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Эволюция лошади
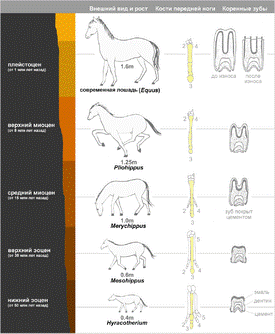
Эволюция лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоев осадочных отложений. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении.
Эволюция лошади достаточно полно прослеживается в Северной Америке. Древнейший представитель лошадиных — гиракотерий (Hyracotherium, также известный как Eohippus) — был размером с лисицу и жил в Северной Америке 54 миллиона лет назад (нижний эоцен), а затем распространился в Европу и в Азию. Это было животное легкого, стройного телосложения, с короткими ногами, но приспособленное к бегу. У него было четыре пальца на передних ногах и три пальца на задних, пальцы располагались практически вертикально. Резцы были маленькие, коренные зубы приплюснуты и покрыты сверху эмалью.
Вероятный путь развития лошадей от гиракотериев к современному виду включает не менее 12 родов и несколько сотен видов. При этом произошли следующие изменения:
· Увеличение размера (от 0.4 м до 1.5 м);
· Удлинение ног и стопы;
· Редукция боковых пальцев;
· Удлинение и утолщение среднего пальца;
· Увеличение ширины резцов;
· Приобретение премолярами (предкоренными зубами) формы, характерной для моляров;
· Удлинение зубов.
· Увеличение высоты коронки коренных зубов.
Судя по ископаемым растениям из различных слоев, болотистая местность, в которой обитал гиракотерий, постепенно становилась суше. Предки лошадей стали полагаться на улучшенный за счет поднятой головы обзор и на высокую скорость бега для спасения от хищников. Если при ходьбе по мягкой болотистой почве большое число пальцев на конечностях было оправдано, то в изменённых условиях жизни для первобытных лошадей стало более выгодным, когда боковые пальцы у них стали постепенно отмирать, а средние — развиваться. Изменение строения зубов объясняется изменением рациона, а именно — переходом с мягкой растительности на траву.
Иногда у лошадей развиваются один или два лишних пальца, чаще всего встречается 2-й (внутренний) палец с вполне развитыми пястными или плюсневыми костями, вполне развитыми суставами пальца и копытом, которое, впрочем, редко касается земли. В случае развития двух лишних пальцев нога получает сходство с ногами предковых видов.
"Микро" и "макро"
Антиэволюционисты, как уже говорилось, в последнее время все чаще соглашаются признать то, что они называют "микроэволюцией" - то есть такие эволюционные изменения, которые можно непосредственно наблюдать в эксперименте или в природе, и которые доказаны настолько убедительно, что даже самые закоренелые антиэволюционисты уже не знают, как это "опровергать", не выставляя себя на посмешище. Сейчас антиэволюционисты, как правило, говорят, что отрицают только то, что они называют "макроэволюцией", то есть крупные эволюционные изменения. В качестве примера "макроэволюционного" события часто называют "выход рыб на сушу", то есть превращение первичноводных позвоночных (рыб) в наземных (четвероногих, или тетрапод).
В науке термином "микроэволюция" принято обозначать эволюционные изменения внутри вида. Акт видообразования считается "пограничным" событием, а появление надвидовых таксонов (новых родов, семейств и т.д.) называют "макроэволюцией". Однако в последние десятилетия эти термины употребляются все реже и реже. Библиотека Pubmed, упоминавшаяся во вводной части, содержит свыше 260,000 статей с ключевым словом "эволюция", но всего-навсего 193 статьи со словом "макроэволюция" и 358 статей со словом "микроэволюция" (по состоянию на 4 марта 2010 г.) Почему же эти термины выходят их моды? Дело в том, что большинство биологов обоснованно считает, что микро-и макроэволюция ничем принципиально не отличаются друг от друга. Точнее говоря, макроэволюция - это просто-напросто суммарный результат множества последовательных микроэволюционных событий. Все упирается в масштаб рассмотрения: когда у нас есть детальная информация (лабораторный эксперимент или очень подробный эволюционный ряд в палеонтологической летописи), мы видим, как правило, последовательность микроизменений. Если же мы рассмотрим только начало и конец длинного ряда (как в случае с эволюцией гоминид, см. выше), то, сравнивая их между собой, можно говорить уже о макроизменениях. Именно такая ситуация получается в том случае, когда не найдены промежуточные формы между двумя крупными таксонами: мы видим значительные изменения и говорим о макроэволюционном "событии". Лишь в редких случаях удается в лаборатории или в природе наблюдать нечто похожее на "макроэволюционное" событие (см. раздел "Эво-дево: следы макроэволюции"). Однако такое сверхускоренное возникновение новых видов с измененным планом строения - это скорее исключение, чем правило. В большинстве случаев на крупные "макроэволюционные" изменения в природе уходят даже не миллионы, а десятки миллионов лет, и складываются они из множества последовательных "микроизменений".
Как говорилось в разделе "Наблюдаемая эволюция", ученым удается, несмотря на все трудности и низкие вероятности, реально наблюдать процесс видообразования в природе и эксперименте. Но можем ли мы точно так же пронаблюдать появление нового семейства? По-видимому, не можем, и вовсе не потому, что таких событий в природе не бывает, а просто по определению, в силу того обстоятельства, что никакая новая форма, выведенная в лаборатории или возникшая на наших глазах в природе, не будет признана новым семейством. Биологи не станут выделять ее в новое семейство именно потому, что все произошло так быстро. Нравится нам это или нет, но ранг семейства биологи присваивают группе только в ретроспективе, когда группа уже "набрала силу", накопила запас изменчивости, разделилась на роды и виды, одним словом, "доказала" всей своей эволюционной историей, что заслуживает семейственного ранга. Даже если в результате "макромутации" возникнет необычная форма с измененным планом строения, потребуются миллионы лет, чтобы выяснить, можно ли эту форму считать родоначальником нового семейства, или это был просто бесперспективный уродец, обреченный на вымирание, или аберрантный представитель предкового семейства, от которого так и не произошло ничего принципиально нового.
Поэтому мы можем реально наблюдать "настоящие" макроэволюционные события только в ископаемой летописи, когда удается найти хорошие филогенетические ряды, соединяющие предковый таксон (например, лопастеперых рыб) с таксоном-потомком (например, первыми бесспорными тетраподами), о котором нам уже заранее известно, что от него в будущем действительно произошло что-то большое, важное и принципиально новое (например, все наземные позвоночные).
Рассмотрим эту ситуацию подробнее. Лопастеперые рыбы дали начало первым наземным позвоночным в конце девонского периода. Это "макроэволюционное событие" довольно детально документировано в ископаемой летописи. Переход от рыб к тетраподам растянулся примерно на 20-25 млн лет (385 - 360 млн лет назад) и состоял из множества последовательных мелких шажков, ни один из которых по отдельности никак "не тянет" на статус макроэволюционного.
Легкие (и, соответственно, два круга кровообращения) были, судя по палеонтологическим данным, у большинства или даже у всех девонских лопастеперых рыб. Они сохранились и у некоторых современных рыб. В девоне было понижено содержание кислорода в атмосфере, а доля углекислого газа, наоборот, была повышена по сравнению с современностью. Поэтому в воде, особенно на мелководье, рыбам было трудно дышать одними жабрами. Они заглатывали воздух, как это делают и современные рыбы, и у них образовались впячивания пищевода для удержания пузырей воздуха. Эти впячивания и есть легкие. Они выполняли сразу две функции - примитивных легких и примитивного плавательного пузыря. В дальнейшем у большинства костных рыб эти древние легкие стали отделяться от пищевода и превратились в "настоящий" плавательный пузырь. Генетический анализ показал, что из современных рыб ближайшими родственниками тетрапод являются двоякодышащие. У двоякодышащих рыб есть и легкие, и два круга кровообращения, и трехкамерное сердце - совсем как у амфибий, которые относятся к настоящим тетраподам.
По сути дела, основное, что изменилось при переходе от рыб к тетраподам - это строение конечностей. Но конечности (плавники) девонских лопастеперых рыб не так уж сильно отличались по своему строению от лап примитивных тетрапод. А главное, можно в деталях проследить изменения конечностей в эволюционном ряду, соединяющем рыб и тетрапод. На сегодняшний день этот филогенетический ряд включает около дюжины последовательных переходных форм, из которых можно упомянуть наиболее важные (в порядке убывания рыбьих и накопления тетраподных черт строения): Eusthenopteron, Panderichthys, Tiktaalik, Elpistostege, Livoniana, Elginerpeton, Ventastega, Metaxygnathus, Acanthostega, Ichthyostega, Tulerpeton(жирным шрифтом выделены роды, у которых известно строение конечностей; у остальных конечности сохранились плохо или вовсе пока не найдены). Первый род в этом списке - еще "типичная рыба", последний - бесспорная амфибия. Посередине помещаются все промежуточные этапы этого "макроэволюционного" преобразования.
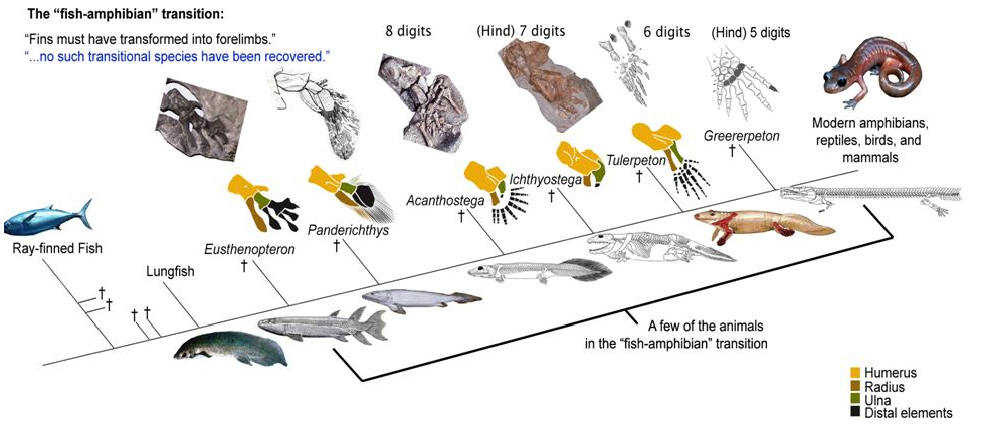
Упрощенная схема перехода от лопастеперых рыб к первым тетраподам. Этот рисунок использовался на "обезьяньем процессе" в Пенсильвании в 2005 г. для иллюстрации ложности заявлений креационистов об отсутствии переходных форм между рыбами и амфибиями (процесс, как известно, выиграли эволюционисты, несмотря на то, что судья Джонс, выносивший вердикт, был консерватором, сторонником Буша и очень религиозным человеком). На этой картинке еще нет тиктаалика, который был найден позже.

Более точные рисунки передних конечностей пандерихта и тиктаалика (предоставил А.Н.Кузнецов).
Уже у Eusthenopteron в переднем плавнике имелась кость, соответствующая плечевой кости тетрапод (показана желтым цветом), и две кости, соответствующие будущим локтевой и лучевой костям (показаны коричневым и зеленым цветом). "Дистальные элементы", соответствующие будущей кисти тетрапод, показаны черным цветом. У пандерихта строение этих дистальных элементов еще оставалось относительно неупорядоченным. У тиктаалика они приобрели чуть более "оформленный" вид, так что становится уже более-менее понятно, откуда взялись у потомков тиктаалика пальцы. Тиктаалик и пандерихт уже могли сгибать свой передний то ли плавник, то ли лапу в суставе, который у их потомков назовут "локтевым". Также у тиктаалика произошла утрата жаберной крышки (которая еще была у пандерихта), что еще на шажок приближает тиктаалика к тетраподам. Утрата жаберной крышки автоматически привела к исчезновению костной связи между поясом передних конечностей и черепом.
У вентастеги и ряда других форм, которые по строению черепа занимают промежуточное положение между тиктааликом и акантостегой, к сожалению, от конечностей почти ничего не сохранилось.
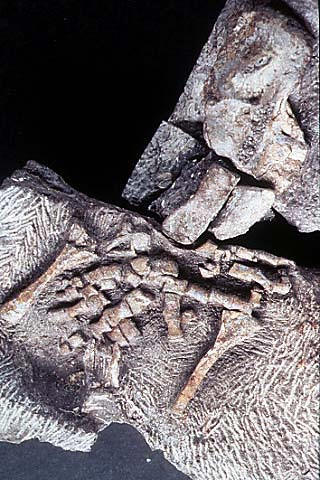
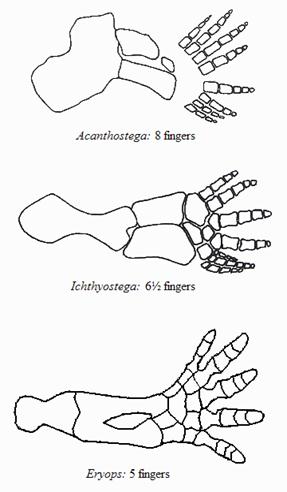
Передняя конечность акантостеги и других ранних тетрапод
Конечности древнейших животных, которых безоговорочно относят уже к тетраподам, то есть к первым амфибиям (акатностега, ихтиостега), представляют собой всего лишь еще один небольшой шажок в сторону дифференциации дистальных частей скелета, т.е. кисти. У акантостеги еще сохранялись внутренние жабры, у ихтиостеги они уже редуцировались (наружные жабры у личинок амфибий сохраняются и сегодня). В дальнейшем у древнейших амфибий происходило постепенное развитие шейного отдела. Совершенно непонятно, где в этом ряду находится искомый великий "макропереход". Сказать, что переход от эустеноптерона к пандерихту и от пандерихта к тиктаалику - это "микро", а вот от тиктаалика к акантостеге - это уже "макро", может только очень предвзятый человек. И, конечно, среди специалистов-палеонтологов нет никого, кому пришло бы в голову это утверждать.
Можно уверенно сказать, что если бы формы, подобные акантостеге и тулерпетону, вымерли, так и не дав начало большой разнообразной группе наземных позвоночных, независимый наблюдатель (скажем, марсианин или какой-нибудь разумный осьминог) никогда бы не выделил их в особый класс, а классифицировал бы как своеобразную тупиковую ветвь полуводных лопастеперых рыб. А осьминоги-креационисты говорили бы: "Это обычная микроэволюция! Рыба осталась рыбой. Покажите нам МАКРОэволюцию!"
Аналогичная ситуация наблюдается сейчас у некоторых современных костистых рыб, которые уже в кайнозое стали предпринимать новые "попытки" освоить сушу, порой весьма успешные. У этих современных рыб предпосылок для выхода на сушу гораздо меньше, чем у их девонских предшественников. Бывшие легкие у них уже "истрачены" на плавательный пузырь, а задние конечности (брюшные плавники) пришли в негодное для превращения в ноги состояние. Тем не менее илистый прыгун ловко ходит по суше, опираясь на грудные плавники и помогая себе хвостом, дышит воздухом при помощи нового, довольно "кустарного" и малоэффективного органа воздушного дыхания, который сформировался у него в ротовой полости. Эта удивительная рыба способна питаться на суше и даже размножается на берегу в "башенках" из глины, на дне которых она устраивает лужицу, где и развивается икра.
Видео: илистые прыгуны настолько уверенно чувствуют себя на суше, что даже охраняют свои участки и устраивают турнирные бои.
(если ролика не видно на странице, его можно посмотреть здесь  )
)
Мы не знаем, дадут ли илистые прыгуны начало новой волне сухопутной экспансии рыб. Может, и дадут, хотя сейчас у них на суше многовато конкурентов. Если все же от них произойдет новая большая сухопутная группа, зоологи будущего придадут ей высокий ранг, а на ее родоночальников - нынешних илистых прыгунов - будут смотреть уже совсем другими глазами. Ну а пока, разумеется, "рыба осталась рыбой".
Илистые прыгуны - далеко не единственные рыбы, которые в наши дни в той или иной мере "воспроизводят" то давнее макроэволюционное событие. Еще один пример - мелководные удильщики антеннарииды. Они не выходят на сушу, однако грудные плавники у них приобрели поразительное сходство с лапами четвероногих. У них есть даже нечто вроде пальчиков!
Видео: мелководный удильщик с лапами, как у наземного позвоночного
(если ролика не видно на странице, его можно посмотреть здесь )
Удильщики при помощи своих лап ползают по дну и держатся за камни, чтобы их не болтало волнами. Как видим, лапы могут быть полезны и под водой. Кстати, большинство палеонтологов считает, что и рассмотренные выше древнейшие тетраподы жили в основном в воде, и лапы у них сформировались преимущественно для подводных нужд (хотя попутно могли использоваться для переползания в другие водоемы).
Лапы удильщиков внешне и по своим движениям очень похожи на лапы тетрапод, но скелет у них другой. Иными словами, это аналогия, а не гомология (чего и следовало ожидать, учитывая независимое эволюционное происхождение этих лап).
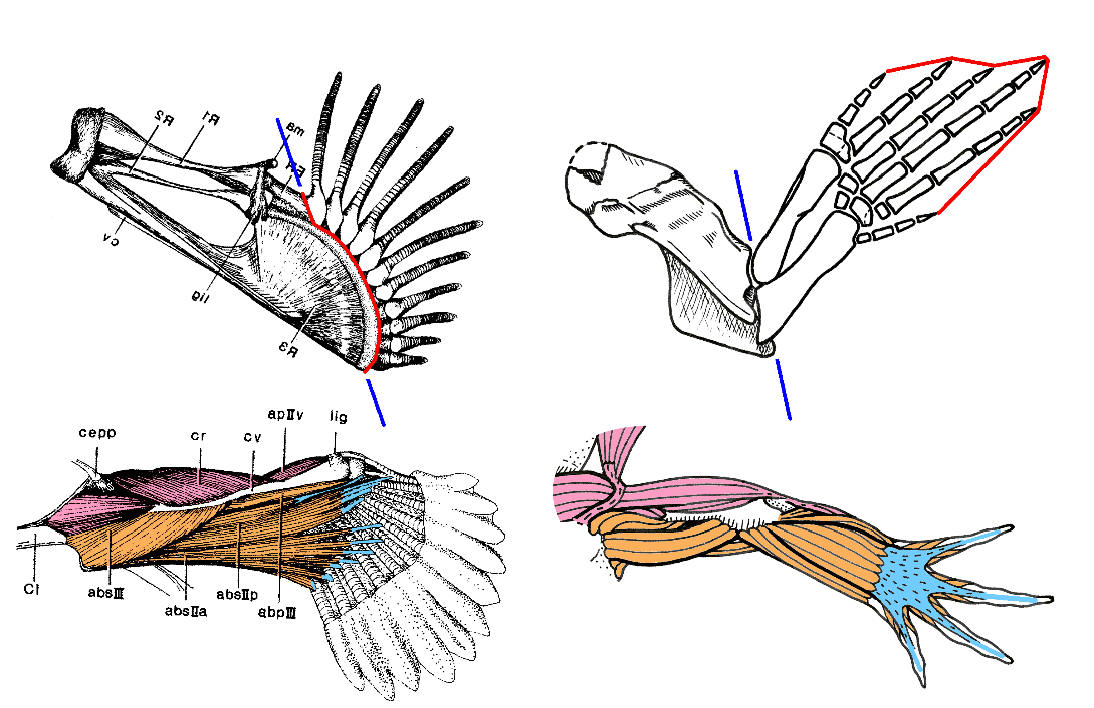
Схема строения передних конечностей мелководного удильщика Antennarius и древней амфибии Tulerpeton. Гомологичные группы мышц показаны одинаковыми цветами. Синей линией показано "главное место сгиба": у тулерпетона это локтевой сустав, а у удильщика - нечто совсем другое, но аналогичное по функции. Изображение предоставил А.Н.Кузнецов.
Можно составить морфологический ряд, восстановив эволюционный путь от обычного рыбьего плавника к "лапе" мелководного удильщика, и тогда креационисты скажут: "вот видите, это обычная микроэволюция, рыба осталось рыбой". Можно не строить ряда, и тогда креационисты скажут: "вот видите, это разумный дизайн". Ученым же эти примеры дают понять, что для рыбы выйти на сушу и стать наземным позвоночным - дело не такое уж хитрое. Причем такое "макро"-превращение в действительности складывается из суммы не очень сложных "микро"-изменений.
Дата добавления: 2015-04-16; просмотров: 176; Мы поможем в написании вашей работы!; Нарушение авторских прав |