КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Оогенез
Тема Строение и развитие половых клеток
1. Оогенез.
2. Сперматогенез.
3. Регуляция сперматогенеза.
4. Отличия оогенеза и сперматогенеза.
Оогенез
Яйцевые клетки образуются в женской половой железе — яичнике (ovarium), расположенной в области малого таза, длиной 2,5-5,5 см, шириной 1,5-3,0 см, толщиной до 2 см, весом 5-8 г. Они проходят долгий путь развития, который начинается в эмбриональном и продолжается в репродуктивном периоде онтогенеза особей женского пола (рис.).
Первичные половые клетки возникают на ранних этапах эмбриогенеза из энтодермальных клеток вегетативного полюса, как, например, у бесхвостых амфибий, или из энтодермальных клеток желточного мешка, как у всех амниот — пресмыкающихся, птиц и млекопитающих. ППК очень рано отличаются от других клеток благодаря крупным размерам и прозрачной цитоплазме. Половые железы в этот момент только начинают формироваться. Экспериментально было показано, что первичные половые клетки мигрируют из места возникновения в развивающиеся гонады и заселяют их. У млекопитающих они перемещаются по дорсальной брыжейке, будучи способными в этот период к амебоидному движению. У птиц миграция происходит пассивно по кровяному руслу. У высших позвоночных не обнаружено веществ, стимулирующих миграцию первичных половых клеток к гонадам. Существует мнение, что первичные половые клетки, оказавшись в каких-либо других участках зародыша, как правило, гибнут, но иногда могут перерождаться в опухоли.
Попав в гонады, первичные половые клетки начинают пролиферировать. Они делятся митозом и называются оогониями. Наступает стадия размножения. У большинства низших позвоночных оогонии сохраняют способность к делению на протяжении всего репродуктивного периода, например, рыбы за один нерест выделяют тысячи яиц, земноводные — сотни (животные с наружным оплодотворением).
Виды, для которых характерно внутреннее оплодотворение, продуцируют половые клетки более экономно. У млекопитающих размножение овогониев протекает только в эмбриональном периоде и к концу внутриутробного развития прекращается. Так, у человека максимальное количество оогониев (6-7 млн) наблюдается у пятимесячного плода. Далее следует массовая дегенерация половых клеток, количество которых у новорожденной девочки составляет около 1 млн, а к семи годам сокращается до 300 тысяч.
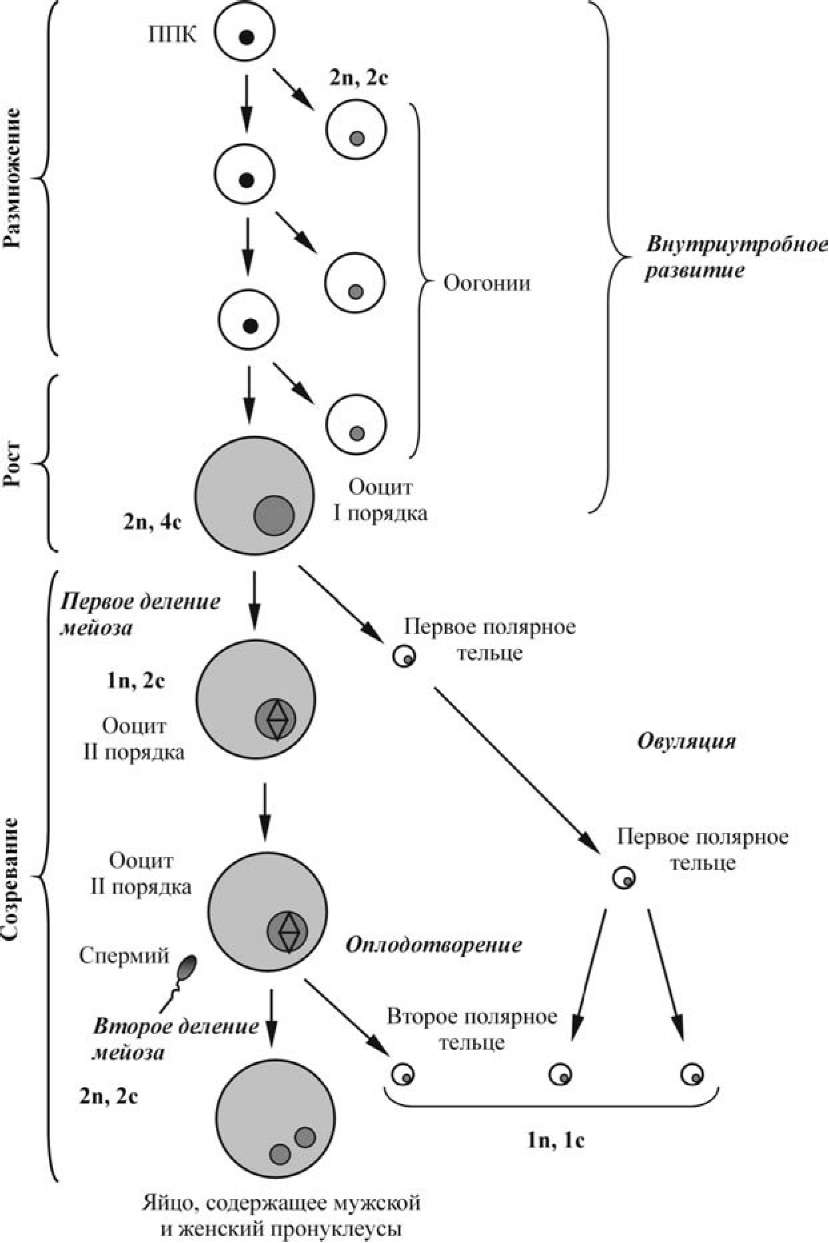 Рис. Схема процесса оогенеза человека
Рис. Схема процесса оогенеза человека
|
Женская половая клетка, прекратившая размножение, называется ооцит I порядка. Начинается свойственный только этой клетке период роста. Он связан с поступлением в яйцеклетку питательных веществ извне и с синтезом их в самой яйцеклетке. Масса и объем яйцеклетки увеличиваются в огромное количество раз (у насекомых — в 90 000 раз, у млекопитающих — более чем в 40 раз).
Рост ооцитов принято разделять на два периода:
• малого, или цитоплазматического, роста (превителлогенез): происходит относительно небольшое пропорциональное увеличение массы ядра и цитоплазмы;
• большого, или трофоплазматического, роста (вителлогенез): резко интенсифицируется рост цитоплазматических компонентов, в ооците откладывается желток.
Весь период превителлогенеза проходит на фоне подготовки ооцита I порядка к последующим делениям созревания (мейоза). Подготовка к первому делению созревания начинается с того, что ооцит вступает в S-период редукционного деления (фаза удвоения ДНК). Затем следует профаза первого деления мейоза, которая продолжается у ооцитов млекопитающих несколько дней.
По достижении фазы диплотены, когда гомологичные хромосомы уже прошли конъюгацию и начали расходиться к противоположным полюсам ядра, наступает стадия диакинеза. На ней дальнейшее течение мейоза сильно замедляется. Остановка мейоза продолжается до достижения особью по- ловозрелости, т. е. длится много лет, применительно к некоторым млекопитающим и человеку. В период диакинеза ядерный материал ооцита не остается инертным: у большинства яйцеклеток он выполняет роль матрицы для синтеза всех видов РНК — информационных, транспортных, матричных и рибосомальных. Все эти типы РНК синтезируются впрок и используются уже оплодотворенной яйцеклеткой. Синтез рРНК связан с уникальным процессом амплификации генов (т. е. временного увеличения числа генов, кодирующих данный вид РНК). Амплификация осуществляется путем избирательного копирования рибосомальных генов, расположенных вдоль нити ДНК. Отделившиеся копии обособляются морфологически в виде ядрышек, которых может насчитываться несколько тысяч.
После созревания ооцита ядрышки выходят в его цитоплазму и там ли- зируются. Синтез рРНК протекает в течение 3-6 месяцев. Низкомолекулярные рРНК и тРНК синтезируются без амплификации — их быстрое накопление обусловлено тем, что кодирующие их гены многократно повторены. Усиливающаяся синтетическая активность нуклеиновых кислот приводит к образованию хромосом типа ламповых щеток, что связано с наличием деспирализованных участков ДНК, на которых идет синтез мРНК. В зрелой яйцеклетке насчитывается до 25-50 тыс. различных типов мРНК.
В период вителлогенеза в ооците I порядка образуется желток, а также жиры и гликоген. Желток — это высокофосфорилированный кристаллический белок. Его количество в клетке строго детерминировано генетически и не зависит от условий питания самки. Вителлогенез может осуществляться за счет синтеза желтка внутри ооцита (эндогенный желток), либо желток синтезируется вне яичника (экзогенный желток). Синтез эндогенного желтка происходит в эндоплазматическом ретикулуме из концевых цистерн аппарата Гольджи. Накопление желтка может идти и в митохондриях, которые при этом перерождаются в желточные гранулы. Большинству видов животных присуще образование экзогенного желтка. Он строится на основе белка, предшественника вителлогенина, поступающего в ооцит извне.
У позвоночных вителлогенин синтезируется в печени матери, транспортируется к содержащему ооцит фолликулу по кровеносным сосудам и поглощается ооцитом путем пиноцитоза. В дальнейшем, при формировании гранул желтка, он распадается на липовителлин и фосфовитин, входящие в состав экзогенного желтка. Синтез вителлогенина клетками печени находится под гормональным контролем. Выделяемый гипоталамусом лю- либерин стимулирует продукцию гонадотропных гормонов гипофиза (ФСГ, ЛГ) в кровь. Под их воздействием клетки фолликула синтезируют в кровоток эстроген. Последний индуцирует, а впоследствии и контролирует, как на уровне транскрипции, так и на уровне трансляции, синтез вителлогенина клетками печени.
Созревание ооцита — это процесс последовательного прохождения двух делений мейоза (делений созревания). При подготовке к первому делению ооцит длительное время находится в фазе диакинеза, когда и происходит его рост и вителлогенез. Начало собственно делений созревания приурочено к достижению самкой половой зрелости и определяется половыми гормонами.
Контроль за процессом созревания яйцеклетки лучше всего изучен у амфибий. У этих животных гонадотропины, находящиеся под контролем гипофиза, действуя на окружающие ооциты фолликулярные клетки, инициируют выделение последними стероидного гормона прогестерона. Подобно другим стероидным гормонам, прогестерон способен диффундировать через плазматические мембраны большинства клеток-мишеней и связываться с внутриклеточными рецепторными белками, регулирующими транскрипцию специфических генов. Однако при созревании ооцита прогестерон, по-видимому, действует иначе. Он связывается с рецепторными белками плазматической мембраны. При этом происходит инактивация плазматической аденилатциклазы, в результате чего снижается концентрация циклического АМФ в цитозоле и соответственно активность цАМФ-зависимой протеинкиназы (А-киназы). Поскольку А-киназа отвечает за фосфорилиро- вание N-концевых участков белков, то ее инактивация приводит к дефосфо- рилированию фактора созревания яйцеклеток (фактор инициации М-фазы, ФИМ), находящегося в цитоплазме. При этом он деблокируется, т. е. переходит в активное состояние.
В норме ФИМ запускает переход из профазы первого деления мейоза в метафазу второго деления. Зрелые ооциты задерживаются на стадии мета- фазы II, когда уровень ФИМ высок. Инактивация А-киназы инициирует активацию небольших количеств ФИМ, которые, в свою очередь, активируют новые порции ФИМ (положительная обратная связь). Замечательное свойство ФИМ — его способность к автокаталитическому саморазмножению, т. е. он может фосфорилировать сам себя и, следовательно, активировать. Фактор созревания вызывает разрушение оболочки ядра ооцита, разрушение ядрышек и миграцию хромосом к будущему анимальному полюсу, где и произойдут деления созревания.
Основная особенность деления созревания в ооцитах состоит в том, что эти деления резко неравномерны. В результате первого деления созревания половина хромосомного набора выталкивается в очень маленькую клетку — редукционное (полярное или направительное) тельце. Впоследствии эта клетка делится на две столь же малые, и никакого участия в дальнейшем развитии они не принимают. Яйцеклетка после выделения первого редукционного тельца называется ооцитом II порядка.
 Рис. 8. Ооцит II порядка после овуляции
(по Алмазову, Сутулову, 1978): 1 — первое редукционное тельце; 2 — веретено второго деления созревания; 3 — блестящая зона; 4 — фолликулярные клетки
Рис. 8. Ооцит II порядка после овуляции
(по Алмазову, Сутулову, 1978): 1 — первое редукционное тельце; 2 — веретено второго деления созревания; 3 — блестящая зона; 4 — фолликулярные клетки
|
Второе деление созревания осуществляется путем выделения второго редукционного тельца таких же размеров, как и первое. После его выделения ооцит II порядка превращается в зрелое яйцо (рис. 8).
Количество яиц, созревающих одновременно, редко достигает 15, обычно их бывает меньше, иногда только одно (человек). У большинства животных течение мейоза останавливается на некотором этапе созревания (блок мейоза), а для дальнейшего его протекания требуется оплодотворение яйцеклетки сперматозоидом (исключение составляют морские ежи и некоторые кишечнополостные).
Различают три типа блока мейоза (именно на этом этапе происходит овуляция яйцеклетки):
• на стадии диакинеза (губки, моллюски, отдельные представители плоских, круглых, кольчатых червей, млекопитающие: собака, лиса, лошадь);
• метафазы 1-го деления созревания (губки, немертины, кольчатые черви, насекомые);
• метафазы 2-го деления созревания (хордовые; у летучих мышей блок мейоза наступает в анафазе 2-го деления созревания).
Дата добавления: 2015-02-09; просмотров: 1118; Мы поможем в написании вашей работы!; Нарушение авторских прав |
| <== предыдущая лекция | | | следующая лекция ==> |
| ПРАВИЛА ПОДГОТОВКИ ПАЦИЕНТА И СБОРА МАТЕРИАЛА ДЛЯ БАКТЕРИОСКОПИЧЕСКОГО ИССЛЕДОВАНИЯ | | |