КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Проникновение сперматозоида в яйцеклетку
Вслед за узнаванием спермия желточной (прозрачной) оболочкой яйца происходит лизис части этой оболочки в области головки спермия, в результате чего плазматические мембраны спермия и яйца сливаются. Поверхность яйца покрыта микроворсинками, контакт между сперматозоидом и яйцом вызывает полимеризацию актина и увеличение размеров микроворсинок, которые формируют воспринимающий бугорок.
Временно существующий воспринимающий бугорок в значительной степени гомологичен акросомному выросту. У морских ежей воспринимающий бугорок может возникать в любой области яйца (рис. 14).
У некоторых амфибий и многих беспозвоночных узнавание спермия и слияние с ним происходит в специализированных областях. Слияние представляет собой активный процесс, часто опосредуемый специфическими «фузогенными» (fusogenic) белками. Известно, что такие белки, как НА-белок вируса гриппа и F-белок вируса Сендай, способствуют слиянию клеток. Не исключено, что биндин относится к белкам именно такого типа.
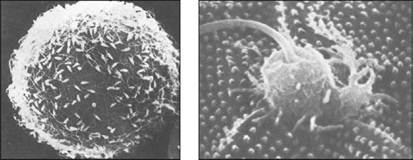 Рис. 14. Оплодотворение у морского ежа (по Гилберту, 1993): А — спермии, прикрепленные к желточной оболочке яйца; Б — спермий вовлекается в яйцо
Рис. 14. Оплодотворение у морского ежа (по Гилберту, 1993): А — спермии, прикрепленные к желточной оболочке яйца; Б — спермий вовлекается в яйцо
|
Предотвращение полиспермии. При нормальном моноспермном оплодотворении гаплоидные ядра спермия и яйца соединяются, образуя диплоидное ядро зиготы, при этом восстанавливается присущее виду число хромосом. Полиспермия (проникновение множества спермиев) приводит у большинства животных к гибельным последствиям. Например, у морских ежей при оплодотворении двумя спермиями возникает триплоидное ядро, в котором каждая хромосома представлена тремя, а не двумя копиями. Вместо нормального процесса — разделения хромосом с помощью биполярного веретена между двумя дочерними клетками — происходит распределение триплоидного набора между четырьмя клетками. В результате одни клетки получают лишние копии некоторых хромосом, тогда как у других эти хромосомы отсутствуют. Такие клетки либо погибают, либо развиваются атипично.
Существуют механизмы защиты яйцеклетки от полиспермии.
Быстрый блок полиспермии. Сразу после контакта первого спермия с плазматической мембраной яйца она (мембрана) должна утрачивать способность сливаться с плазматической мембраной спермия. Быстрый блок полиспермии достигает этой цели путем изменения электрического потенциала плазматической мембраны яйца (рис. 15).
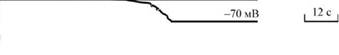 Рис. 15. Мембранный потенциал в яйце морского ежа до и после оплодотворения (по Гилберту, 1993)
Рис. 15. Мембранный потенциал в яйце морского ежа до и после оплодотворения (по Гилберту, 1993)
|
| В нормальном состоянии на мембране ооцита поддерживается потенциал, равный -70 мВ. В течение 0,1 с после прикрепления первого спермия мембрана яйца деполяризуется, при этом мембранный потенциал достигает положительных величин (+20 мВ). Это является следствием резкого изменения Na+ проницаемости мембраны, в результате чего положительно заряженные ионы натрия поступают внутрь яйцеклетки (по градиенту концентрации), «уничтожая» отрицательный заряд внутренней стороны цитоплазматической мембраны, тем самым уменьшая исходную (-70 мВ) разность потенциалов. Открытие натриевых каналов в яйце индуцируется, по-видимому, прикреплением к нему спермия. В настоящее время из спермиев морского ежа выделен акросомный белок, способный открывать натриевые каналы в неоплодотворенном яйце. |
Медленный блок полиспермии. Быстрый блок полиспермии действует недолго. Мембранный потенциал яйца морского ежа остается положительным только около 1 мин. Это кратковременное смещение потенциала может быть недостаточно для предотвращения полиспермии. Удаление избыточных спермиев, прикрепленных к желточной оболочке, осуществляется посредством кортикальной реакции. Этот более медленный блок полиспермии начинает действовать примерно через 1 мин после прикрепления спермия к яйцу.
В яйце морского ежа непосредственно под плазматической мембраной располагаются около 15 000 кортикальных гранул. После контакта спермия с яйцом в присутствии Ca2+ эти кортикальные гранулы сливаются с плазматической мембраной и выделяют содержимое в область между плазматической мембраной и желточной оболочкой (рис. 16).
Белки, связывающие желточную оболочку и поверхность яйца, разрушаются высвобождающимися пептидазами:
• вителлиновая деламиназа — отделяет желточную оболочку от цито- плазматической мембраны яйцеклетки;
• сперморецепторная гидролаза — освобождает поверхность яйца от осевших на желточной оболочке сперматозоидов, лизируя сайты их соединения.
Выделяемые мукополисахариды и осмотически активный гликопротеид создают осмотический градиент, обусловливающий поступление воды из цитоплазмы яйца в пространство между желточной оболочкой и плазматической мембраной (перивителлиновое пространство). В результате объем яйцеклетки несколько уменьшается, желточная оболочка отделяется от поверхности яйца и с этого момента называется оболочкой оплодотворения. В процессе кортикальной реакции она претерпевает ряд изменений:
• воздействие протеаз приводит к изменению свойств биндиновых рецепторов или к их отделению вместе с прикрепленными к ним спермиями;
• ферменты из кортикальных гранул разрушают связи желточной оболочки
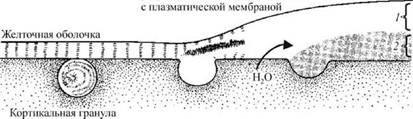 Рис. 16. Формирование оболочки оплодотворения (по Гилберту, 1993): 1 — оболочка оплодотворения; 2 — гиалиновый слой
Рис. 16. Формирование оболочки оплодотворения (по Гилберту, 1993): 1 — оболочка оплодотворения; 2 — гиалиновый слой
|
• пероксидаза, содержащаяся в кортикальных гранулах, вызывает затвердевание оболочки путем образования поперечных связей между остатками тирозина соседствующих белков.
Образование оболочки оплодотворения начинается в месте проникновения спермия (примерно через 20 с после прикрепления) и отсюда распространяется по всей поверхности яйца, завершаясь к концу первой минуты после прикрепления оплодотворяющего спермия. Одновременно происходит выделение гиалина — белка, запасенного в кортикальных гранулах, который образует сплошной слой вокруг яйца. Плазматическая мембрана взаимодействует с этим белком, и гиалиновый слой поддерживает бластомеры в период дробления.
У млекопитающих кортикальная реакция не вызывает образования оболочки оплодотворения, но результат ее оказывается таким же, как и у морских ежей: рецепторы спермиев изменяются, и они больше не удерживают сперматозоиды на поверхности яйца. Этот процесс изменения свойств рецепторов спермиев носит название реакции прозрачной оболочки или реакции zona.
У млекопитающих частота полиспермии бывает минимальной благодаря малому числу спермиев, достигающих места, где происходит оплодотворение. Механизмы защиты от полиспермии, наблюдаемой при оплодотворении богатых желтком яиц (некоторые птицы, рептилии, амфибии), когда в цитоплазму проникает несколько сперматозоидов, неясны. При этом все спермии, за исключением одного, разрушаются, после того как женский пронуклеус сольется с мужским.
Механизм кортикальной реакции сходен с механизмом акросомной реакции. Прикрепление спермия к плазматической мембране яйца через активацию находящегося в мембране G-белка стимулирует активность другого мембраносвязанного фермента — фосфолипазы С. Она, в свою очередь, расщепляет находящийся здесь же фосфатидилинозитол-4,5-бифосфат на диацилглицерол — ДАГ (остается связанным с мембраной), и инозитолтрифосфат — ИТФ (диффундирует в цитоплазму).
Диацилглицерол посредством протеинкиназы С активирует Na+/Н+-ан- типортер, что приводит к увеличению внутриклеточного pH (натрий входит в клетку, а протон ее покидает) и как следствие активации белкового синтеза, репликации ДНК и перемещения морфогенетических детерминант в цитоплазме (при участии Ca2+). ИТФ вызывает высвобождение Ca2+ из внутриклеточных депо (эндоплазматического ретикулума), что приводит к экзоцитозу кортикальных гранул и включению медленного блока полиспермии, а также образованию гиалинового слоя вокруг яйца.
Другим Ca2+-зависимым эффектом является активация НАД-киназы, которая катализирует превращение НАД в НАДФ+, что в конечном итоге обеспечивает синтез новых липидных компонентов плазматической мембраны, необходимых для последующего протекания процесса дробления.
Дата добавления: 2015-02-09; просмотров: 434; Мы поможем в написании вашей работы!; Нарушение авторских прав |