КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Физическая природа следов памяти
Синаптическая гипотеза. По мнению Хебба (Hebb, 1974), различия между кратковременной и долговременной памятью обусловлены главным образом различиями в структурах нервных сетей.
Сенсорная и кратковременная память,1 согласно гипотезе Хебба, обусловлена повторной циркуляцией (реверберацией) сигналов по многочисленным нервным путям, образующим замкнутые цепи. Поскольку сигналы при этом постоянно возвращаются к одним и тем же пунктам, возбуждение нейронных контуров может некоторое время поддерживаться, и одновременно может происходить посылка импульсов к другим центрам или по двигательным путям (рис. 8.16).
Что касается долговременной памяти, то она обусловлена, по мнению Хебба, длительным изменением синаптических связей, возникающим в результате повторной циркуляции импульсов. Благодаря этому создается все более и более прочный след, лежащий в основе памяти. Однако для того, чтобы этот след мог закрепиться, соответствующие контуры должны некоторое время оставаться неактивными. Этот период, длящийся от 15 минут до часа, называют периодом консолидации, и в это время происходит закрепление новых знаний или навыков. Именно поэтому после сотрясения мозга человек не может вспомнить о тех событиях, которые произошли непосредственно перед травмой, а остальные воспоминания нарушаются тем меньше, чем они дальше во времени от момента травмы.
Биохимические гипотезы. Известно, что видовая генетическая память записана на молекулах ДНК (дезоксирибонуклеиновой кислоты). ДНК содержится в ядрах всех клеток тела и представляет собой набор генов. На основе информации, содержащейся в ДНК, образуется другое вещество - РНК (рибонуклеиновая кислота), которая управляет функциями клетки, определяя синтез специфических белков. Белки играют первостепенную роль как в построении тканей, так и в различных функциях организма (см. приложение А).
Исследование химических изменений. Естественным образом возник вопрос: не может ли РНК -вещество, столь близкое к ДНК, -быть тем ключевым элементом, от которого зависит образование белков, специфических для разных видов научения.
На этот вопрос в 50-х годах пытался ответить пионер биохимических исследований в области памяти - шведский ученый Хиден (Hyden, 1969). Для этого он вырабатывал у крыс и мышей различные навыки, при которых изменялось их обычное поведение. Например, он заставлял животное получать пишу, балансируя на проволоке или действуя с помощью не той лапки, которой оно пользуется обычно. Хиден обнаружил, что после такого изменения поведенческих реакций не только
26 437
402 Глава 8
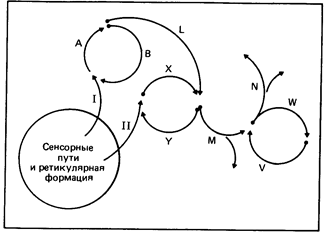
Рис. 8.16. Схема, иллюстрирующая синаптическую теорию памяти. Информация, поступающая от рецепторов, может более или менее прямым путем направляться к центрам, ответственным за немедленное принятие решения, либо передаваться к двигательным центрам по путям A, L, М и N или же X, М и N. Однако одновременно эта информация может циркулировать по кругам, в которых, например, структура А вызывает возбуждение В, а та в свою очередь - опять возбуждение А и т. д. (либо по аналогичному кругу из структур Х или Y). От того, как долго будет сохраняться возбуждение в путях I или II, а также от уровня активации организма зависит, перейдет ли след из кратковременной памяти в долговременную.
увеличивалось общее количество РНК в мозгу, но и отмечались также сдвиги в ее качественном составе. Значит, при научении действительно происходят изменения на уровне молекул, как количественные, так и качественные. Хиден даже выделил особую молекулу, которую он назвал S100 и которая, по его мнению, как раз и была «молекулой памяти», ответственной за освоение новых навыков. Но хотя эти результаты и были многократно воспроизведены, они не дали ответа на вопрос о том, действительно ли новые молекулы специфически связаны именно с научением или же все эти количественные и качественные изменения просто сопровождают активацию мозга.
Изучение действия химических ингибиторов памяти. Медикам хорошо известно, что антибиотики подавляют синтез белков микроорганизмами. Это обусловлено ингибирующим действием антибиотиков на образование РНК. Аналогично действует и рибонуклеаза – фермент, разрушающий РНК и препятствующий ее образованию.
Поэтому интересно было проверить, нельзя ли, вводя такого рода ингибиторы в мозг, уничтожить какие-либо приобретенные реакции или помешать формированию новых.
Подобные исследования предпринимались многими учеными. Некоторые из них (например, Агранофф) проверили эту гипотезу на рыбках, которые были обучены избегать одного из отсеков аквариума. Джон
Память, мышление и общение
(John, 1967) вводил рибонуклеазу в мозг кошки, у которой была выработана зрительная дифференцировка. Флекснер (Flexner, 1967) вводил антибиотик в мозг мыши, обученной избегать одну из ветвей Т-образного лабиринта. Результаты всех этих опытов были примерно одинаковыми. Введение подобных веществ в мозг после обучения действительно приводило к «стиранию» следов памяти, и животное должно было обучаться заново. В то же время такие вещества не влияли ни на кратковременную память, если вводились сразу же после обучения, ни на долговременную, если их вводили спустя длительное время после выработки навыка. Значит, стиратели следов, несомненно, действуют во время периода консолидации, о котором мы говорили выше. Однако достаточно ли этого, чтобы можно было говорить о молекулярном кодировании, которое будто бы и подавляется подобными веществами?
Эксперименты с «переносом молекул памяти». Увлечение идеями молекулярного кодирования памяти подтолкнуло некоторых ученых к попыткам проверить, нельзя ли осуществить биохимическую передачу каких-то навыков от одних животных другим. В 60-х годах Мак-Коннел и его сотрудники одними из первых проделали подобные опыты на планариях. Планарии -маленькие плоские черви, о которых мы уже говорили в первой главе, -это одни из самых простых животных, у которых имеется подобие мозга.
Исследователи вырабатывали у планарий условную реакцию на включение лампочки, которое сопровождалось электрическим ударом. Поскольку планарий -это животные, пожирающие себе подобных, исследователи растирали в порошок обученных планарий и скармливали необученным. Оказалось, что после этого у таких необученных планарий условные реакции на свет формировались гораздо быстрее, чем у их собратьев, которым скармливали таких же необученных червей.
Вдохновленные этими результатами, Мак-Коннел и его сотрудники сумели даже выделить из планарий-доноров РНК и ввести ее планариям-реципиентам. При этом тоже был достигнут эффект переноса навыка. По-видимому, сходные результаты были получены и на крысах (McConnel et al., 1970).
Однако столь многообещающие, казалось бы, результаты были встречены многими учеными с недоверием. Представление о «передаче знаний с помощью молекул» вызвало ряд критических замечаний. Указывалось, например, что «формирование условных реакций» на свет могло быть просто сенсибилизацией к этому раздражителю, усиленной в результате поедания ткани уже сенсибилизированной особи.
Действительно, когда у планарий были выработаны более сложные навыки (например, выбор пути в Y-образном лабиринте), эффект переноса уже не проявлялся. Значит, маловероятно, чтобыРНК сама по себе играла здесь ведущую роль.
Унгар (Ungar, 1970)-венгерский ученый, работавший в США, -исследовал выработку избегания определенных мест у крыс и мышей. У этих животных существует врожденная инстинктивная склонность прятаться
404 Глава 8
в темных уголках, однако каждый раз, когда они забегали в затемненный ящик, они получали удар электрическим током. Довольно быстро у них выработалась настоящая боязнь темных мест. После этого Унгар вводил экстракты растертого мозга таких животных необученным реципиентам; в результате оказалось, что животные после этого проводили гораздо меньше времени в темном ящике, чем их собратья, которым был введен гомогенат от необученных доноров. Более того, гомогенизировав мозг сотен обученных крыс, Унгар выделил из него в чистом виде пептид, который назвал скотофобином (дословно: «вызывающий страх темноты»). В дальнейшем он сумел искусственно синтезировать этот пептид и получил с его помощью аналогичные результаты.
Однако в данном случае, по-видимому, речь тоже не шла о «молекулах памяти» в том смысле, в каком о них говорил Мак-Коннел. По мнению Унгара, подобные молекулы действуют скорее всего на уровне синапсов, где они играют роль «указателей», способствующих циркуляции нервных импульсов именно по тем путям, которые необходимы для консолидации нейронных цепей.
Новые нейрофизиологические подходы. Поскольку во всех этих работах исследовались довольно сложные виды научения, в связи с их результатами возникали серьезные вопросы, а интерпретация оказывалась уязвимой для критики.
В 70-х годах некоторые ученые, например Кэндел (Kandel, 1976), пошли по другому пути: они решили тщательно изучить такие простые виды научения, как привыкание (габитуация).
Кэндел ставил свои опыты на аплизии (морском зайце) - крупном моллюске до 30 см длиной (рис. 8.17). У аплизии имеется сифон, с помощью которого она втягивает воду и пропускает ее под мантией; при этом вода фильтруется и из нее извлекаются мелкие организмы, служащие для аплизии кормом. Прикосновение к сифону вызывает реакцию втягивания жабры. Но если это раздражение повторяется, то наступает привыкание и рефлекторное втягивание жабры сначала ослабевает, а затем и вовсе исчезает.
Выбор аплизии был обусловлен тем, что нервная система этого животного функционирует примерно так же, как у позвоночных, и, кроме того, у него относительно мало нервных волокон, идущих к ганглиям. Вдобавок эти волокна довольно толстые (до 1 мм) и поэтому нетрудно установить, откуда они идут и куда приходят. В такие волокна Кэндел мог легко вводить электроды, с помощью которых можно было регистрировать импульсы, идущие от рецепторов к ганглиям или от ганглиев к мышцам. Такая методика позволила Кэнделу тщательно изучить, что происходит в синапсах тех нейронов, которые участвуют в процессе привыкания. Он, в частности, установил, что ослабление двигательной реакции в случае привыкания обусловлено уменьшением количества медиатора, выбрасываемого в синаптическую щель, и соответствующим уменьшением частоты импульсного разряда постсинаптического нейрона.
Память, мышление и общение 405
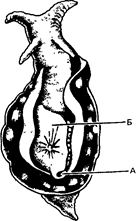
Рис. 8.17. Аплизия, или морской заяц. Этот крупный моллюск всасывает воду через сифон (Л), а затем пропускает ее под мантией (Б), где из воды отцеживаются мелкие организмы, которые служат для аплизии пищей.
Разумеется, подобные исследования еще очень далеки от тех тончайших методов регистрации, которые потребуются при изучении нервной системы человека. Однако они позволяют получать четкие и однозначные данные при изучении клеточных аспектов кратковременной памяти у примитивных животных.
Кроме того, результаты этих исследований позволяют наметить первые точки соприкосновения между поисками энграммы, как представлял их себе Лэшли, синаптической теорией и исследованием механизмов облегчения и торможения передачи нервных сигналов с помощью химических медиаторов (см. приложение А).
Литература
Общие работы
Chapouthier G., 1980. "Les bases physiologiques de 1'apprentissage et de la memoire",
dans Psychophysiologie, Paris, Ed. Etudes Vivantes. Hebb D.O., 1974. Psychologie, science moderne, Montreal, Ed. HRW. Kandel E., 1976. Cellular basis of behavior. An introduction to behavioral neurobiology,
San Francisco, Freeman. Kimble D.F. (sous la direction de), 1965. The Anatomy of Memory, Palo Alto, Calif.,
Science and Behavior Books. Lashley K., 1929. Brain Mechanisms and intelligence, Chicago, University of Chicago
Press. McConnel J. V. (1962). "Memory transfer through Cannibalism in Planarians", Journal
of Neuropsychiatry, n° 3 (suppl. 1), p. 542-548. McConnel J. V., Shigehisa Т., Salive H., 1970. "Attempts to transfer approach and
avoidance responses byRNA injections in rats", in:K.H. Pribram etD.E.
Broadbendt (Eds.), Biology of Memory, New York, Academic Press. Penfieid W., 1975. The Mystery of the Mind: A Critical Study of Consciousness and the
Human Brain, Princeton, N.J., Princeton University Press.
406 Глава 8
Pribram К., 1969. Languages of the Brain, Englewood Ciffs, N. J., Prentice-Hall. Ungar G. (sous la direction de), 1970. Molecular Mechanisms in Memory and Learning. Plenum Press.
Цитированная литература
Bronckart J.P., 1977. Theories du langage, Bruxelles, Pierre Mardaga. Bruner J.S., Goodnow J.J., Austin G.A., 1956. A Study of Thinking, New York, John
Wiley and Sons. Bruner J.S., 1966."On cognitive growth: II", in: J. S. Bruner,R.R. Olver, P.M.
Greenfield, 1966, Studies in cognitive growth, New York, Wiley. Bower G.H., Trabasso T. (1963). "Reversals prior to solution in concept identification",
Journal of experimental Psychology, n° 66, p. 409-418. Chomsky N.(1965). Aspects de la theorie de la syntaxe, Paris, Seuil, 1971. Chomsky N. (1968). Le langage et la pensee, Paris, Payot, 1970. Collins A.M., Quillian M.R. (1969). "Retrieval time from semantic memory". Journal of
Verbal Learning and Verbal Behavior, n° 8, p. 240-247. Curtiss S., 1977. Genie: A psycholinguistic study of a modern-day "wild child", New
York, Academic Press. Davis F.C. (1932). "The functional significance of imagery differences". Journal of
experimental Psychology, n° 15, p. 630-661. Ehrlich S., Tulving E. (1973-1976). "La memoire semantique", Bulletin de Psychologie,
n° 29 (numero special).
Ehrlich S., 1975. Apprentissage et memoire chez 1'homme, Paris,PUF.Fouts R.S. (1973). "Acquisition and testing ofgestual signs in four young chimpanzees",
Science, n°, 180, p. 978-980. Gardner R., Gardner B. (1969). "Teaching sign Language to a Chimpanzee", Science,
n° 165, p. 664-672.
Hayes C., 1951. The ape in our house, New York, Harper and Row. Hering E., 1912. Theory of Light Sensation, Boston, Houghton Mifflin. Hultsch D.F. (1971). "Organisation and memory in adulthood". Human Development,
n° 14, p. 12-29.
Inglis J., Ankus M.N., Sykes D.H. (1968). "Age-related differences in learning and short-term memory from childhood to the senium", Human Development, n° 11, p. 42-52. lingram D. (1975). "Surface contrasts in children's speech", Journal of Child Language,
n° 2, 287-292.
Kellogg W.N., Keliogg L.A., 1933. The Ape and the Child, New York, McGraw-Hill. Kosslyn S.M. (1978). "Measuring the visual angle of the mind's eye", Cognitive
Psychology, n° 10, p. 356-389.
Leippe M.R., Wells G.L., Ostrom T.M. (1978). "Crime seriousness as a determinant of accuracy in eyewitness identification". Journal of Applied Psychology, n° 63, p. 345-351.
Lenneberg E.H„ 1967. Biological foundations of language. New York, John Wiley. Levine M., 1975. Hypothesis testing: A cognitive theory of learning, Hillsdale, N.J.,
Lawrence Eribaum Associates. Lieury A., 1975. La memorie, Bruxelles, Dessart. Lieury A., 1980. Les precedes mnemotechniques, Bruxelles, P. Mardaga ed. Loftus E.F. (1979). "The malleability of human memory", American Scientist, n° 67,
p. 321-320.
Mervis C.V., Rosch E. (1981). 'Categorization of natural objects", Annual Review of Psychology, n° 32, p. 89-115.
Память, мышление и общение
Miller G. A. (1956). "The magical number seven, plus or minus two: Some limits of our
capacity for processing information", Psychological Review, n° 63, p. 81-97. Morgan С. Т., 1974. Introduction a la psychologie, Montreal, McGraw-Hill. Mowrer, 1960. Learning theory and behavior. New York, John Wiley. Patterson F.G. (1978). "The gestures of a gorilla: Langauge acquisition in another
pongid". Brain and Language, n° 5, p. 72-97. Peterson L.R., Peterson M.J. (1959). "Short-term retention of individual verbal items",
Journal of Experimental Psychology, n° 58, p. 193-198. Piaget J; Inhelder В., 1966. La psychologie de 1'enfant, Paris, P.U.F. Piaget J., Inhelder В., 1966. L'image mentale chez 1'enfant-Etude sur le development des
representations imagees, Paris, P. U. F. Premack D. (1976). "Language and intelligence in ape and man", American Scientist,
n° 64, p. 674-683.
Richelle M., 1971. L'acquisition du langage, Bruxelles, Dessart et Mardaga. Rumbaugh D.M., 1977. Language Learning by a Chimpanzee: the Lana project, New
York, Academic Press. Sapir E., 1921. Language, New York, Harcourt. Seron X., 1979. Aphasie et neuropsychologie, Bruxelles, P. Mardaga ed. Sperling G. (I960). "The information available in brief visual perceptions", Psychological
Monographs, n° 74, (11, integral n° 498). Terrace H. S., 1980. Nim: un chimpanze qui a appris le langage gestuel, Bruxelles,
P. Mardaga ed.
Tulving E., Pearlslone Z. (1966). "Availability versus accessibility of information in memory for words". Journal of Verbal Learning and Verbal Behavior, n° 5, p. 381-391. Tulving E., 1972. "Episodic and semantic memory", in: E. Tulving et W. Donaldson
(Eds), Organization of memory. New York, Academic Press. Wallas G., 1926. The art of thought, New York, Harcourt, Brace and World. Whorf B.L., 1956. Language, thought, and reality, New York, John Wiley. Zeigarnik B. V. (1927). "On finished and unfinished tasks", in: W. D. Ellis (Ed.), A Source Book of Gestalt Psychology, London, Kegan Paul, French, Trubner, 1938, p. 300-302, 312-314.
Дата добавления: 2015-02-10; просмотров: 235; Мы поможем в написании вашей работы!; Нарушение авторских прав |