КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Нециклический транспорт электронов и нециклическое фотофосфорилирование
В ФСII димер П680, поглотив энергию, эквивалентную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние П680*, отдает 2 электрона феофетину (Фф).
От Фф электроны, теряя энергию, последовательно передаются на пластохиноны ФС II – QA и QB, на пул липидорастворимых молекул пластохинона (PQ), переносящих через липидную фазу мембраны электроны и протоны, на железосерный белок FeSR и цитохром f цитохромного комплекса b6–f, восстанавливая Cu-содержащий белок пластоцианин (Пц).
Вакантные места («дырки») в П680 заполняются двумя электронами из содержащего Мn переносчика электронов Z, который в свою очередь восстанавливается с участием системы S. Окисленный белковый комплекс S связывает воду и восстанавливается за счет электронов воды. Для осуществления этой реакции в белковом комплексе S необходимы Мn, Сl– и Са2 + .
При возбуждении П700 в реакционном центре ФС I энергией, эквивалентной 2 квантам длинноволнового красного света, 2 электрона захватываются мономерной формой хлорофилла a (A1) и затем последовательно передаются переносчикам электронов А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и ферредоксин:NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза восстанавливает NADP + .
На вакантные места в П+700 переходят электроны с Пц, и нециклическая цепь переноса электронов таким образом замыкается.
Энергия, освобождающаяся при движении электронов от П680 (Е'о = – 0,8 В) до П700 (Е'о = +0,4 В), используется для синтеза АТР из ADP и неорганического фосфата (фотофосфорилирование).
Таким образом, нециклический транспорт электронов сопровождается выделением кислорода и восстановлением НАДФ+ и сопряжен с синтезом АТФ (нециклическое фотофосфорилирование).
Циклический транспорт электронов и циклическое фотофосфорилирование.
Циклический транспорт электронов осуществляется отдельно в ФС I или ФСII.
Циклический транспорт в ФС I включает перенос электронов от восстановленого ферредоксина(Фдвост.) обратно к окисленному П+700. В этом случае возбужденные молекулы П700 последовательно отдают электроны на А1, А2, АВ, Фд, PQ, цитохром b6, FeSR, цитохром f, Пц и, наконец, П700. При этом NADP+ не восстанавливается. Освобождающаяся энергия используется для фосфорилирования ADP.
Циклический поток электронов с участием ФСI сопряжен с с синтезом АТФ (циклическое фотофосфорилирование) и может обеспечивать дополнительный синтез АТФ, необходимый для ассимиляции углерода. Он может быть альтернативным путем использования энергии света при его высоких интенсивностях.
Циклический транспорт электронов в ФС II связан с обратным переносом электронов от восстановленных первичных хинонов QA и QB к окисленному пигменту П+680. При этом участвуют цитохром b559, β-каротин и сопровождающие молекулы хлорофилла а РЦ ФСII.
Является альтернативным путем использования энергии света. Он активируется в условиях, когда интенсивность света превышает возможности ЭТЦ утилизировать его энергию или при повреждении водоокисляющей системы хлоропластов.
Псевдоциклический транспорт электронов – перенос электронов от воды на кислород – впервые был исследован А. Меллером (1951) и назван реакция Меллера.
Восстановление кислорода может происходить как в ФС I, так и в ФС II. При этом поглощение кислорода может компенсировать его выделение в ходе окисления Н2О. В результате единственным продуктом этого процесса, как и в случае циклического транспорта электронов, будут АТФ, синтезируемая при псевдоциклическом фотофосфорилированиии.
4. Электронтранспортная цепь митохондрий растительной клетки: структурная организация, основные компоненты, их окислительно-восстановительные потенциалы. Пять состояний дыхательной цепи переноса электронов (по Чансу)
Дыхательная электронтранспортная цепь (ЭТЦ), локализована во внутренней мембране митохондрий и служит для передачи электронов от восстановленных субстратов на кислород, что сопровождается трансмембранным переносом ионов Н+. Таким образом, ЭТЦ митохондрий (как и тилакоидов) выполняет функцию окислительно-восстановительной Н+-помпы.
Б. Чанс и др. (США) в 50-х годах, используя значения окислительно-восстановительных потенциалов известных в то время переносчиков е−, спектрофотометрические данные о временной последовательности их восстановления и результаты ингибиторного анализа, расположили компоненты ЭТЦ митохондрий в следующем порядке:


 Сукцинат ФАД 1/2О2+2Н+
Сукцинат ФАД 1/2О2+2Н+
Е°' -0,045
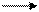

 НАДН ФМН Q Цит Цит Цит Цит
НАДН ФМН Q Цит Цит Цит Цит
b c1 c a, a3
Е°', В  Н2О
Н2О
-0,320 -0,300 +0,070 +0,080 +0,235 +0,250 0,300
Пара электронов от НАДH или сукцината передается по ЭТЦ до кислорода, который, восстанавливаясь и присоединяя два протона, образует воду.
Основным фактором, определяющим способность какого-либо соединения присоединять или отдавать электроны, является соотношение величин окислительных потенциалов данного соединения и соединения, с которым оно взаимодействует. Непосредственное взаимодействие соединений, имеющих различные потенциалы, осуществляется самопроизвольно тем успешнее, чем меньше различия между их потенциалами. В дыхательной цепи перенос электрона от НАДН (Е° = -0,32) на О2 (Е° = +0,82) идет через цепь переносчиков по градиенту, т. е. от соединений с более отрицательным к соединениям с более положительным редокс-потенциалом. По величине положительного потенциала более всего к потенциалу кислорода приближается цитохромоксидаза. К потенциалу дыхательного субстрата наиболее близки потенциалы, свойственные пиридиннуклеотидам, и т. д.
Согласно современным данным дыхательная цепь митохондрий включает в себя четыре основных мультиэнзимных комплекса и два небольших по молекулярной массе компонента — убихинон и цитохром с.
Комплекс I осуществляет перенос электронов от NADH к убихинону Q. Его субстратом служат молекулы внутримитохондриального NADH, восстанавливающиеся в цикле Кребса. В состав комплекса входит флавиновая FMN-зависимая NADH-убихинон-оксидоредуктаза, содержащая три железосерных центра.
Комплекс II катализирует окисление сукцината убихиноном. Эту функцию осуществляют флавиновая (FAD-зависимая) сукцинат убихинон-оксидоредуктаза, в состав которой также входят три железосерных центра.
Комплекс III переносит электроны от восстановленного убихинона к цитохрому с, т. е. функционирует как убихинол-цитохром с-оксидоредуктаза. В своем составе он содержит цитохромы b556 и b560, цитохром с1 и железосерный белок Риске. В присутствии убихинона комплекс III осуществляет активный трансмембранный перенос протонов.
В терминальном комплексе IV электроны переносятся от цитохрома с к кислороду, т. е. этот комплекс является цитохром с-кислород-оксидоредуктазой (цитохромоксидазой). В его состав входят четыре редокс-компонента: цитохромы а и а3 и два атома меди. Цитохром а3 и СuВ способны взаимодействовать с О2, на который передаются электроны с цитохрома а−CuА Транспорт электронов через комплекс IV сопряжен с активным транспортом ионов Н +.
В результате изучения пространственного расположения компонентов ЭТЦ во внутренней мембране митохондрий показано, что комплексы I, III и IV пересекают мембрану. Именно в этих трех участках окислительные процессы в ЭТЦ сопрягаются с синтезом АТР. Передача 2е− от сукцината на убихинон в комплексе II не сопровождается трансмембранным переносом протонов.
Пять состояний дыхательной цепи переноса электронов (по Чансу)
Согласно Чансу митохондрии могут находиться в пяти состояниях:
I) дыхание осуществляется только за счет имеющихся в интактных митохондриях эндогенных субстратов и систем окисления и фосфолирования;
II) наступает после дополнительного внесения субстрата фосфорилирования (АДФ). Лимитирующим фактором в этих условиях является субстрат. Все элементы ЭТЦ в данных условиях находятся в окисленном состоянии;
III) возникает после совместного внесения АДФ и субстрата окисления, что сопровождается резким активированием процесса. Лимитировать дыхание может только проницаемость митохондриальных мембран и скорость окисления;
IV) наступает вследствие исчерпания всей внесенной АДФ и ее превращения в АТФ. Наблюдающееся в этих условиях торможение дыхания и соответствует тому, что называют дыхательным контролем.
Дополнительное внесение АДФ при наличии кислорода обусловливает новое активирование дыхания и фосфорилирования;
V) соответствует полному поглощению кислорода и возникновению условии, необходимых для анаэробного процесса, в результате чего все компоненты ЭТЦ находятся в восстановленной форме.
Дата добавления: 2015-04-15; просмотров: 415; Мы поможем в написании вашей работы!; Нарушение авторских прав |