КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
БУФЕРНЫЕ СИСТЕМЫ
Буферные системы — это биологические жидкости организма. Их защитная роль в поддержании нормального рН крови чрезвычайно велика.
Любая буферная система представляет собой смесь слабой кислоты и ее соли, образованной сильным основанием. Попадание в плазму сильной кислоты вызывает реакцию буферных систем, в результате которой сильная кислота превращается в слабую. То же происходит и при действии на биологические жидкости сильного основания, которое после взаимодействия с буферными системами превращается в слабое основание. В результате указанных процессов изменения рН либо не наступают, либо являются минимальными.
Гидрокарбонаты обеспечивают 53 % буферной способности крови, 47 % ее относятся к негидрокарбонатным системам: гемоглобиновой (35 %), протеиновой (7 %) и фосфорной (5 %). Кровь составляет только 1/5 общей буферной емкости организма.
Гидрокарбонатная система. Происхождение гидрокарбонатной системы тесно связано с метаболизмом органического углерода, поскольку конечным продуктом его является СО2 или НСО3. Гидрокарбонатный буфер является главной и единственной буферной системой интерстициальной жидкости. Образующаяся в клетках СО2 вступает в реакцию с водой, в результате чего получается угольная кислота, которая диссоциирует на ионы Н+ и НСО3. При определенных обстоятельствах (сдвиг реакции вправо или влево) будет преобладать тот или иной тип реакции:

Выделение СО2 происходит через легкие, ионы H+ и НСО3 выделяются через почки. Избыточное образование СО2 ведет к усиленной элиминации его через легкие, и равновесие восстанавливается.
Гемоглобиновая система. Буферное действие молекулы гемоглобина происходит за счет имидазольной группы гистидина. Диссоциация этой имидазольной группы зависит от насыщения кислородом: оксигенированный гемоглобин (HbО2), являясь более сильной кислотой, чем деоксигенированный, отдает больше H+. Благодаря этому облегчается связывание СО2 в тканевых капиллярах и освобождение его в легочных, и транспорт СО2 происходит при меньших сдвигах рН, чем при постоянном SO2 (эффект Кристиансена — Дугласа — Холдена).
Протеиновая система. Белки плазмы, являясь амфолитами, в крови обладают свойствами кислот. Они составляют наибольшую часть пула анионов плазмы. Изменение содержания альбуминов, протеинов и аномальных белков плазмы оказывает существенное влияние на величину так называемой анионной разницы.
Фосфатная система. Эта система — первичный и вторичный фосфат (Н3РО- и НРО-) имеет значение для внутриклеточного пространства и при забуферивании мочи. Анионы РО-, как и органические кислоты, относятся к группе неизменяемых анионов, играющих важную роль в изменениях метаболического компонента КОС.
КОС и рН крови зависят от четырех факторов: продукции фиксированных (нелетучих) кислот, буферирования фиксированных кислот, элиминации фиксированных кислот, элиминации летучей (углеродной) кислоты.
Роль легких в регуляции КОС. Организм — своего рода открытая система, в которой метаболические процессы определяют интенсивность обмена энергии с внешней средой. Процессы образования СО2 в тканях, взаимодействия с гидрокарбонатным буфером и выделения СО2 легкими находятся в состоянии биологического равновесия.
ЗАКЛАДКА (ПРОВЕРИТЬ ФОРМУЛУ)
В нормальных условиях источником энергии является аэробный гликолиз:С6Н.зОб -> 6СО2 + Н2О.
ПосколькуСО2 легко диффундирует через полупроницаемые мембраны, РСО2 приблизительно одинаково во всех жидкостных средах организма.
За сутки в клетках образуется от 13 000 до 20 000 ммоль СО2. Нормально функционирующие легкие могут элиминировать любое количество образующейся СО2 в тканях. Равновесие достигается в том случае, если количество образованной СО2 равно количеству элиминированной.
Роль почек в регуляции КОС. Почки непосредственно экскретируют Н+ из кислой среды и НСО3 из щелочной. При снижении уровня гидрокарбоната в плазме проксимальные канальцы реабсорбируют его до концентрации 25 ммоль/л. в результате чего гидрокарбонатный буфер восстанавливается (механизм щажения оснований путем ионогенеза). На каждый восстановленный НСО3 с мочой экскретируется один H+. Значительное количество Н+ выделяется почками в связанной форме — в норме за сутки через почки выделяется 100—200 ммоль H+. Биологическое равновесие достигается в том случае, если количество образованных в тканях кислот равно количеству выделенных.
Ликвидация протонов с помощью фосфатной системы происходит путем образования дегидрофосфата из монофосфата:
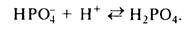
Действие этого механизма непродолжительное. При его истощении реабсорбция Na+ и НСО3 осуществляется за счет аммониогенеза. При увеличении количества ионов во внеклеточной жидкости в клетках почечных канальцев образуется аммиак путем дезаминирования некоторых аминокислот. Аммиак легко диффундирует в канальцевую мочу, где соединяется с ионами Н+:
NН3 + Н+ =NH4+
Образованные ионы аммония не могут вновь проникнуть через клеточную мембрану. Они присоединяют Сl- и в виде NН4Cl выводятся с мочой. Ионы Na+ освобожденные от Сl-, в клетках почечных канальцев соединяются с освобожденными от H+ ионами НСО3 и в виде гидрокарбоната поступают в венозную кровь.
Реакции буферных систем крови при увеличении в ней концентрации протонов происходят обычно до включения почечного механизма компенсации. Почки лишь постепенно увеличивают выделение кислот, иногда в течение нескольких дней, что и является причиной медленной компенсации ацидоза.
Основные компоненты КОС:
• рН — отрицательный десятичный логарифм концентрации H+, величина активной реакции крови. В норме рН артериальной крови 7,4 (7,36—7,44), венозной крови 7,37 (7,32—7,42). Внутриклеточное значение рН 6,8—7,0;
• РСO2 — респираторный компонент КОС. В норме PCO2 артериальной крови равно 40 (36—44) мм рт.ст., венозной крови — 46 (42—55) мм рт.ст.;
• НСО3 — содержание аниона гидрокарбоната (бикарбоната) в плазме крови. В норме составляет 22—25 ммоль/л в артериальной и 25— 28 ммоль/л в венозной крови. Увеличение содержания НСО3 указывает на метаболический алкалоз, а снижение — на метаболический ацидоз;
• BE — избыток или дефицит оснований, отражающий состояние метаболического компонента КОС. Тесно коррелирует с уровнем НСО3. В норме нет ни избытка, ни дефицита оснований и BE равен нулю с колебаниями от -2,3 до +2,3 ммоль/л. Увеличение оснований в алкализированной крови соответствует понятию избытка оснований и обозначается символом BE со знаком «+». Например, величина ВЕ=+5 ммоль/л указывает на то, что в 1 л крови пациента имеется избыток оснований, равный 5 ммоль или дефицит ионов Н+ равный также 5 ммоль. Уменьшение оснований в ацидотической крови соответствует понятию дефицита оснований и обозначается символом BE со знаком «—»;
• ВВ — буферные основания, сумма всех буферных анионов: гидрокарбоната, фосфата, белков и гемоглобина в миллимолях на 1 л крови при 37 °С и РСO2 40 мм рт.ст. В норме ВВ составляет 50 (40— 60) ммоль/л, отражает только метаболические влияния.
Величина рН крови зависит от двух величин: РСО3 крови (респираторный компонент) и содержания оснований в ней (BE, НСО3), что составляет метаболический (нереспираторный) компонент. Последний остается постоянным при острых сдвигах РСО2, хотя хронические изменения РСО2 связаны с компенсаторными изменениями этих показателей.
Дата добавления: 2015-04-15; просмотров: 262; Мы поможем в написании вашей работы!; Нарушение авторских прав |