КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Микробиологи утверждают: многоклеточность — сплошное жульничество
«Чувство кворума»: принятие коллективных решений в макро- и микромире)
Очень важно понять, что эволюционируют не взрослые организмы, а генетические программы их индивидуального развития (онтогенеза). В ходе эволюции меняются не сами морфологические признаки, а генетически обусловленные алгоритмы их формирования в онтогенезе. Данные эмбриологии свидетельствуют о том, что алгоритм развития каждого вида живых организмов является модификацией алгоритмов развития его предков. Генетическая программа онтогенеза многоклеточных животных, как ни странно, сама по себе намного проще (содержит меньше информации), чем получающийся на ее основе взрослый организм. Новая информация "самозарождается" в ходе онтогенеза (в этом суть процесса самоорганизации). Чтобы понять эволюцию многоклеточных, необходимо четко представлять себе основные принципы онтогенеза (в доходчивой форме они описаны здесь). Возможные (допустимые, "разрешенные") пути эволюционных преобразований определяются не столько строением взрослого организма, сколько устройством его онтогенеза.
Понимание процессов самоорганизации имеет ключевое значение для понимания эволюции. Все мы неоднократно наблюдали такой яркий пример самоорганизации, как образование поразительно красивых и сложных морозных узоров на стекле. При этом у нас на глазах из "простого" (из хаотического движения молекул водяного пара) рождается "сложное" - изысканные ледяные картины, достойные кисти лучших художников. Мы понимаем, что эти узоры рисует не дед Мороз, они образуются сами. Но где они "закодированы", где хранится чертеж или программа, на основе которой они формируются? По-видимому, эта программа "закодирована" в физических свойствах молекулы воды, от которых зависит ход кристаллизации, а также в условиях среды (температура воздуха, текстура стекла и т.п.). Примерно таким же образом и программа развития организма закодирована в геноме и окружающей среде (нужно помнить, что строение взрослого организма зависит отнюдь не только от генов, но и от условий среды, в которой происходит развитие). Мы смиряемся с тем очевидным фактом, что и в ходе образования ледяных узоров, и в ходе развития организма происходит самопроизвольное образование новой информации (на основе простой программы самоорганизуется нечто значительно более сложное). Мы не считаем это чудом (хотя на чудо это очень похоже!) и не привлекаем сверхъестественные силы для объяснения каждой снежинки и каждого онтогенеза. Все сказанное в полной мере приложимо и к эволюционному развитию жизни. Наследственная изменчивость и дифференциальное размножение - это просто еще один механизм самоорганизации, благодаря которому из первоначальных простейших существ сами собой с неизбежностью развиваются те самые "endless forms most beautiful and most wonderful" (бесчисленные прекрасные и удивительные формы), которые населяют нашу планету. Подобно ледяным узорам, "закодированным" в молекуле воды и текстуре стекла, все это поразительное многообразие жизни было "закодировано" в самых первых живых существах, в их ДНК или РНК, и в условиях среды, где им предстояло эволюционировать.
Индивидуальное развитие многоклеточных организмов далеко не всегда, но довольно часто повторяет ("рекапитулирует") отдельные этапы его эволюционной истории. Так получается потому, что мутации, меняющие ранние этапы развития, чаще оказываются вредными и реже - полезными, чем мутации, затрагивающие более поздние этапы. Чем раньше в ходе эволюции появился признак, тем больше вероятность, что какие-то другие жизненно важные признаки организма уже успели стать зависимыми от него ("подогнались", "притерлись" к нему под действием отбора), и поэтому он не может быть изменен без ущерба для целого. Напротив, новые признаки часто очень пластичны, потому что все прочие признаки в ходе индивидуального развития формируются независимо от этого нового признака. Поэтому часто самый простой способ для эволюции "создать" новый признак состоит в том, чтобы "пристроить" программу его формирования в самый конец уже имеющейся, устоявшейся программы.
Таким образом, эволюционная теория предсказывает, что в эмбриональном развитии иногда должны воспроизводиться этапы эволюционной истории, а ранние стадии развития зародышей должны быть в тенденции более похожими друг на друга, чем поздние. Эти предсказания хорошо подтверждаются фактами. Антиэволюционизм, напротив, ничего такого не предсказывает, потому что "разумный дизайнер" имел полную возможность разработать уникальную, неповторимую программу индивидуального развития для каждого вида в отдельности. Однако, сравнивая между собой эмбриональное развитие разных животных, мы видим не набор уникальных программ, а поразительное и очень глубокое сходство. Например, даже очень непохожие друг на друга животные проходят практически одинаковые ранние стадии развития: зигота (оплодотворенное яйцо), бластула, гаструла и др. Как и в морфологии (см. раздел "Морфологические доказательства"), в индивидуальном развитии организмов мы видим бесчисленные и несомненные следы "происхождения путем модификации", а вовсе не множества независимых актов творения.
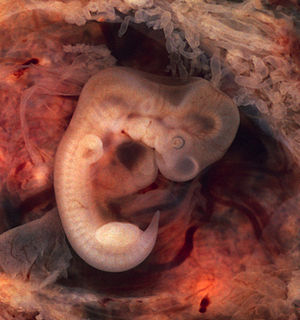
Месячный эмбрион человека при внематочной беременности. Видны зачатки жаберных дуг и хвост.
У всех позвоночных животных наблюдается значительное сходство зародышей на ранних стадиях развития: форма тела, зачатки жаберных дуг, хвост, один круг кровообращения и т. д. (закон зародышевого сходства Карла Максимовича Бэра). Однако по мере развития сходство между зародышами различных систематических групп постепенно стирается и начинают преобладать черты, свойственные их классам, семействам, родам, и, наконец, видам.
Эволюционные изменения могут касаться всех фаз онтогенеза, то есть могут приводить к изменениям не только зрелых организмов, но и эмбрионов, даже на первых этапах развития. Тем не менее, более ранние фазы развития должны отличаться большим консерватизмом, чем более поздние, так как изменения на более ранних этапах развития в свою очередь должны привести к большим изменениям в процессе дальнейшего развития. Например, изменение типа дробления вызовет изменения в процессе гаструляции, равно как и во всех следующих стадиях. Поэтому изменения (мутации), проявляющиеся на ранних этапах, чаще оказываются летальными, чем изменения, касающиеся более поздних периодов онтогенеза.
Таким образом, ранние стадии развития изменяются относительно редко, а значит, сравнительное изучение эмбрионов разных видов может быть источником сведений о степени эволюционного родства.
В 1837 году эмбриолог Карл Райхерт выяснил, из каких зародышевых структур развиваются квадратная и сочленовная кости в челюсти рептилий. У зародышей млекопитающих обнаружены те же структуры, но они развиваются в молоточек и наковальню среднего уха (см.: Н.Н.Иорданский. Эволюция жизни. Глава 3). Палеонтологическая летопись также подтверждает происхождение частей уха млекопитающих из костей челюсти рептилий.
Изучение развития зародышей современных организмов помогает реконструировать ход эволюции и даже позволяет предсказывать, какие палеонтологические находки могут быть сделаны в будущем. Так, у предков зверей — зверозубых рептилий — была только одна слуховая косточка (стремечко), а молоточек (=сочленовная кость) и наковальня (=квадратная кость) образовывали челюстной сустав. То же самое наблюдается и у самых примитивных древних млекопитающих. У современных млекопитающих в ходе эмбрионального развития эти косточки сначала, как и у далеких предков, прилегают к нижней челюсти (на этом этапе они неотличимы от сочленовной и квадратной костей эмбрионов рептилий), а затем отделяются от нее. Отделение происходит в два этапа: сначала слуховые косточки отделяются от боковой поверхности нижней челюсти, но остаются связаны с ней спереди посредством меккелева хряща (этот хрящ, кстати, представляет собой остаток первичной, изначальной нижней челюсти позвоночных; у акул, например, вся нижняя челюсть образована меккелевым хрящом). На втором этапе исчезает и это переднее соединение: меккелев хрящ у взрослых млекопитающих рассасывается. Таким образом, эмбриональное развитие млекопитающих подтверждает выводы сравнительной анатомии о происхождении молоточка и наковальни млекопитающих от костей челюстного сустава рептилий.
На основании этих фактов свыше 30 лет назад была выдвинута гипотеза, согласно которой в эволюции среднего уха млекопитающих должна была быть промежуточная стадия — примерно такая же, как в эмбриональном развитии современных зверей (Allin, E. F. 1975. Evolution of the mammalian middle ear. // J. Morphol. 147, 403–438). Эта гипотеза блестяще подтвердилась в 2007 году, когда в Китае был найден скелет млекопитающего, жившего 125 млн лет назад (в раннемеловую эпоху) и получившего название яноконодон (Yanoconodon allini). Молоточек и наковальня у яноконодона уже отделены от нижней челюсти с боков, но еще прикреплены к ней спереди посредством окостеневшего меккелева хряща ( см.: Новая палеонтологическая находка проливает свет на раннюю эволюцию млекопитающих)..
Есть многие другие примеры того, как эволюционная история организма временно проявляется в ходе его развития. У эмбрионов млекопитающих на ранних стадиях есть зачатки жаберных дуг и жаберных мешков, неотличимые по строению от соответствующих зачатков у зародышей водных позвоночных. Это объясняется тем, что предки млекопитающих жили в воде и дышали жабрами. Разумеется, жаберные мешки зародышей млекопитающих в ходе развития превращаются не в жабры, а в структуры, возникшие в ходе эволюции из жаберных щелей или стенок жаберных карманов, такие как евстахиевы трубы, среднее ухо, миндалины, паращитовидные железы и тимус. При этом отчасти сохраняется относительное расположение нервов и кровеносных сосудов, подходивших к жаберным дугам у рыб. Именно поэтому, например, возвратный гортанный нерв у млекопитающих от мозга идет вниз по шее, огибает аорту и возвращается вверх к гортани (см. раздел "Морфологические доказательства").

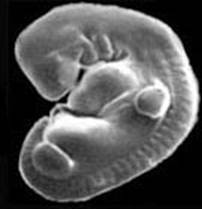
Эмбрион кота (слева) и человека (справа) на стадии, когда хорошо видны жаберные дуги, хвост, зачатки передних и задних конечностей. С сайта talkreason.org
У эмбрионов многих видов змей и безногих ящериц (например, веретеницы ломкой) развиваются зачатки конечностей, но затем они рассасываются.[см. статью о редукции конечностей у рептилий] Аналогично, у китов и дельфинов нет задних конечностей, но у эмбрионов китообразных начинают расти задние ноги, развиваются кости, нервы, сосуды, а затем все эти ткани рассасываются (Amasaki, H., et al. Developmental changes of the fore- and hind-limbs in the fetuses of the southern minke whale, Balaenoptera acutorostrata; Sedmera, D., et al. On the development of Cetacean extremities: I. Hind limb rudimentation in the Spotted dolphin Stenella attenuata)
Дарвин в качестве примера приводил наличие зубов у зародышей усатых китов (Дарвин, 1859. Глава XIV. Рудиментарные, атрофированные и абортивные органы.)

Эмбрион дельфина Stenella attenuata. Видны зачатки передних (f) и задних (h) конечностей. Из первых разовьются плавники, вторые исчезнут. С сайта talkreason.org
У человеческих эмбрионов на ранних этапах развития (4-5 недель) есть заметный хвост с 10-12 развивающимися хвостовыми позвонками, длина которого составляет более 10 % длины эмбриона (Fallon, J. F., Simandl, B. K. Evidence of a role for cell death in the disappearance of the embryonic human tail). Развиваются не только позвонки, но и спинной мозг, и ряд других структур. К восьмой неделе шесть крайних позвонков отмирают (в результате смерти клеток). 5-й и 4-й хвостовые позвонки уменьшаются, прочие структуры также отмирают и уменьшаются . Макрофаги утилизируют разрушающиеся клетки (Sapunar, D., et al. Morphological diversity of dying cells during regression of the human tail). У взрослых людей остается 4 хвостовых позвонка, которые образуют копчик.
Современные сумчатые и плацентарные млекопитающие живородящи, но их предки были яйцекладущими (подобно современным ехиднам и утконосам). У плацентарных в эмбриональном развитии не осталось явных следов скорлупы или специальных приспособлений для ее пробивания, однако в ходе развития зародышей сумчатых на короткое время возникает и потом рассасывается яичная скорлупа, а у некоторых новорожденных сумчатых (коала, бандикуты и др.) имеется рудиментарный яйцевой зуб (Tyndale-Biscoe, H. and Renfree, M. 1987. Reproductive Physiology of Marsupials).
Любимым аргументом антиэволюционистов является тот факт, что Эрнст Геккель, более ста лет назад нарисовавший свой знаменитый рисунок, демонстрирующий сходство эмбрионов разных классов позвоночных, на самом деле сильно идеализировал реальную картину (что, впрочем, в те времена не считалось таким уж страшным прегрешением - ведь он рисовал просто схему, а схемы всегда идеализированы). Но какое нам дело сейчас, спустя полтора века, до рисунков Геккеля? Они - не более чем исторический курьез, и их неточность никоим образом не влияет на силу и достоверность эмбриологических доказательств эволюции. Посмотрите хотя бы выше на фотографии эмбрионов человека, кота и дельфина. Впрочем, и научная недобросовестность Геккеля вовсе не является доказанным фактом, о чем свидетельствую данные, приведенные в статье R.J.Richards, 2009. Haeckel’s embryos: fraud not proven.
Дополнительный пример 1. Подкласс двусторонне-симметричных "неправильных" морских ежей (Irregularia) произошел от обычных "правильных" радиально-симметричных морских ежей (подкласс Regularia) в юрском периоде. Эволюционный переход от правильных ежей к неправильным, начавшийся около 170 и закончившийся около 145 млн лет назад, детально документирован в палеонтологической летописи (см. А.Н.Соловьев, А.В.Марков. Ранние этапы эволюции неправильных морских ежей). При этом у всех современных неправильных ежей личинка сначала превращается в маленького радиально-симметричного ежика, строение которого во всех основных чертах воспроизводит морфологию правильных морских ежей. В ходе дальнейшего развития постепенно развивается двусторонняя симметрия и все основные отличительные признаки подкласса Irregularia.

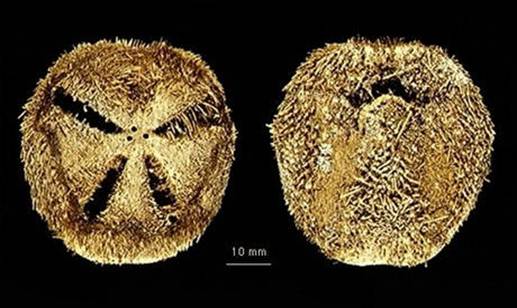
"Новорожденные" неправильные морские ежики рода Abatus (слева) имеют радиальную симметрию и по своему строению в целом соответствуют диагнозу подкласса Regularia. Взрослые Abatus (справа), как и все представители подкласса Irregularia, имеют двустроннюю симметрию.

Ископаемый морской еж Pygaster (юрский период) - одна из переходных форм между правильными и неправильными морскими ежами. Преобладает радиальная симметрия, но перипрокт (анальное отверстие) уже сильно смещен назад (на фото - вниз), что придает ежу элементы двустронней симметрии. В индивидуальном развитии современных неправильных морских ежей приобретение двусторонней симметрии тоже начинается со смещения перипрокта из центра верхней поверхности в заднем направлении.
Дополнительный пример 2. У взрослых камбалообразных рыб оба глаза находятся на одной стороне головы, а весь череп сильно искривлен и искорежен. У предков камбал, естественно, голова была симметричной, как у всех нормальных рыб. Поэтому неудивительно, что мальки камбал тоже имеют симметричную голову, которая в ходе роста постепенно теряет симметрию, искажается, и один глаз переползает на другую сторону головы. Недавно обнаружены ископаемые переходные формы, доказывающие возможность постепенного эволюционного перехода от "обычных" рыб к камбалообразным. См. в заметке Ископаемые рыбы в очередной раз подтвердили правоту Дарвина
См. также замечательную подборку фактов на англ. языке с сайта talkorigins.org.
Дата добавления: 2015-04-16; просмотров: 352; Мы поможем в написании вашей работы!; Нарушение авторских прав |