КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ГЛАВА 3. ФАКТОРЫ ЭВОЛЮЦИИ
В предыдущих главах мы рассмотрели материальные предпосылки эволюции (наследственную изменчивость) и биологические макросистемы - виды и популяции, являющиеся структурными единицами эволюционного процесса. Теперь мы можем перейти к рассмотрению движущих сил эволюции, важнейшей из которых является естественный отбор.
Естественный отбор
Как уже было отмечено, мутационная изменчивость необходима, но сама по себе еще недостаточна для осуществления приспособительной и прогрессивной эволюции. Приспособленность к конкретным условиям существования - важнейшая предпосылка выживания и нормальной жизнедеятельности организмов - формируется только благодаря действию естественного отбора. Естественный отбор является творческой силой эволюции, направляющей (векторизующей) эволюционный процесс и интегрирующей отдельные изменения в адаптации. В этом современные представления вполне соответствуют взглядам Дарвина. Однако следует отметить, что отбор не требует перенаселения как обязательного условия, обеспечивающего его эффективность. Борьба за существование, понимаемая в том широком и метафорическом смысле, который вкладывал в него сам Дарвин, означает всю совокупность взаимоотношений организма и внешней среды и, следовательно, должна иметь место и без перенаселения. Естественный отбор поэтому должен действовать и в ситуациях, далеких от перенаселения.
Генетическая сущность отбора заключается в неслучайном (избирательном) выживании изменчивых генотипов и в избирательном участии разных генотипов в передаче генов последующим поколениям. Следовательно, естественный отбор имеет два существенно различающихся аспекта: отбор на выживание, определяющий дифференцированное выживание разных особей, и репродуктивный отбор (не путать с половым отбором), обеспечивающий различную степень участия разных особей в размножении. Эти два аспекта отбора могут не совпадать - например, стерильный гибрид может обладать повышенной жизнеспособностью.
Различные факторы внешней среды, воздействующие на данный вид организмов и требующие определенной приспособительной реакции организмов, представляют собой отдельные силы отбора (или селективные силы). Общее воздействие отбора на данный вид организмов, иногда называемое давлением отбора или селективным давлением, складывается как результат интеграции отдельных селективных сил. Взаимодействие последних друг с другом может оказывать существенное влияние на общий эффект отбора.
Если разные селективные силы имеют несовпадающие направления, общий итог их взаимодействия получает характер эволюционного компромисса, подразумевающего формирование под контролем отбора такого фенотипа, который обеспечивает скорее некоторое оптимальное соотношение разных адаптации, чем достижение максимального совершенства одной из них.
Необходимо отметить, что до сих пор определенное распространение (особенно среди критиков дарвинизма) имеют упрощенные и, по существу, неверные представления о естественном отборе, которые ведут к серьезным ошибкам в понимании механизмов эволюционного процесса. Прежде всего неверно представление о естественном отборе как изолированном действии отдельных селективных сил на отдельные признаки организма, т. е. как отборе отдельных признаков (и тем более - отдельных аллелей). Темпы эволюции, рассчитанные на основе этих представлений, оказываются гораздо более низкими, чем наблюдаемые реально поданным палеонтологии темпы эволюции многих филетических линий, а плата за отбор - непомерно высокой. Напомним, что отбору непосредственно подвергаются не отдельные гены, а определенные фенотипы, формирующиеся в онтогенезе на основе соответствующих генотипов при модифицирующем влиянии внешних условий. Следовательно, отбираются целые генотипы, характеризующиеся такой нормой реакции, которая наиболее благоприятна в данных условиях среды по всей совокупности признаков. Благодаря блочно-иерархическому принципу организации генома отбор может действовать на целостные генетические комплексы (супергены), что на несколько порядков ускоряет эволюционный процесс.
Поскольку всякий генотип характеризуется специфическим спектром мутационных изменений (т. е. обладает способностью с некоторой частотой давать определенные мутации), естественный отбор, благоприятствуя конкретным генотипам, тем самым благоприятствует и соответствующему направлению и частоте возникновения наследственных вариаций. Таким образом, если отбор благоприятствует каким-то фенотипическим признакам, носителем которых является данный генотип, то отбор тем самым влияет и на характер дальнейших эволюционных изменений.
Преобладающие направления изменчивости различных генотипов могут в разной степени соответствовать направлению отбора. В этом случае при наличии двух генотипов, которые являются носителями какого-то нового полезного признака, но различаются своими спектрами изменчивости, отбор, очевидно, будет в большей мере благоприятствовать тому генотипу, возможности изменчивости которого будут ближе к направлению действия отбора. При совпадении преобладающего направления изменчивости и направления отбора приспособительная эволюция пойдет гораздо быстрее, чем при отсутствии такого совпадения. Быстрота возникновения и усовершенствования приспособлений дает очевидное преимущество в борьбе за существование. Таким же образом отбор может воздействовать и на частоту определенных мутаций (через посредство соответствующих генов-мутаторов), поскольку этот показатель существенно влияет на распространенность в популяции благоприятных вариантов.
Следовательно, естественный отбор, выбраковывая одни и сохраняя другие генотипы, тем самым оказывает влияние на направление и частоту возникающих наследственных изменений, что также способствует значительному ускорению эволюционного процесса.
Вторая группа ошибочных представлений о естественном отборе связана с непониманием его творческой роли. Эти представления коренятся в упрощенной концепции отбора, которая на первый взгляд кажется очевидной: отбор понимается как своего рода "сито", устраняющее наименее приспособленные варианты наследственных изменений (мутаций) и сохраняющее более приспособленные. Именно такое понимание отбора характерно, например, для сторонников "нейтральной эволюции" (М. Кимура, С. Оно и др.), подчеркивающих "консервативную природу" отбора и видящих движущую силу эволюции в мутационном процессе (см. гл. 1). Этот очевидный результат действия отбора, несомненно, существует, но отнюдь не исчерпывает его эффект во всей полноте. При концентрации внимания на "консервативной роли" отбора странным образом забывают, что отбор не только выбраковывает неудачные варианты, но и накапливает удачные, формируя из них адаптации любой степени сложности, и преобразует эти адаптации в соответствии с изменениями внешних условий.
Отбор оказывает на организм многостороннее интегрирующее влияние, способствуя оптимизации взаимодействий разных его систем. Приспособительное, формообразовательное и интегрирующее воздействие отбора на организм и обозначается как творческая роль естественного отбора.
Интенсивность действия отбора
Естественный отбор можно представить как вектор, поскольку его, с одной стороны, характеризует величина, которую можно выразить количественно, с другой стороны - направление.
Количественная характеристика действия отбора носит название его интенсивности или эффективности: она представляет собой степень воздействия (давления) отбора на популяцию. Очевидно, давление отбора в разных условиях может быть большим или меньшим. Для количественного выражения интенсивности отбора используют показатель, называемый коэффициентом отбора.
Пусть некоторая популяция включает два типа особей - А и В.
Концентрации особей А и В в популяции соотносятся как СА/СВ=u1.
Если один из этих вариантов имеет преимущество перед другим, измеряемое коэффициентом интенсивности отбора S, то в следующем поколении соотношение концентраций особей А и В в популяции изменится в соответствии с этим коэффициентом:
u2= u1(1-S)
S=(u1-u2)/u1=1-u2/u1
где u2 – соотношение концентрации особей в следующем поколении.
Если u1>u2, то S>0 и особи В имеют какое-то преимущество перед особями А, которое "отмечается" отбором, и концентрация особей В в популяции возрастает от поколения к поколению. Если же u1=u2, то S = 0, следовательно, особи А и В равноценны и соотношение их концентраций в данной популяции не меняется.
Коэффициент интенсивности отбора позволяет сравнивать давление отбора на разные фенотипы. Для полиморфной популяции устанавливают коэффициент отбора всех вариантов по отношению к наиболее приспособленному (т.е. наиболее жизнеспособному и плодовитому в данных условиях) фенотипу. Очевидно, коэффициент отбора представляет собой величину относительную и не может характеризовать действие отбора только на данный фенотип без сравнения с другими вариантами.
Формы отбора
Если интенсивность давления отбора является его количественной характеристикой, то направление естественного отбора определяет качественную специфику его воздействия на популяцию. Выделяют три основных направления действия отбора, о которых обычно говорят как о типах (или формах) естественного отбора.
Стабилизирующий отбор. Если условия внешней среды длительное время остаются стабильными, приспособленность вида к этим условиям постепенно совершенствуется, пока не достигнет определенного оптимума. Совершенство всякого приспособления, разумеется, относительно, обычно возможна и более высокая степень приспособленности к данным условиям. Однако эволюционные возможности всякого конкретного генофонда, приспосабливаемого отбором к данным условиям среды хотя и велики, но ограничены спектром возможных мутационных изменений. Поэтому раньше или позже при постоянстве условий среды и при отсутствии конкуренции с экологически близкими формами генофонд данного вида оптимизируется по отношению к существующим условиям. С этого момента на популяцию начинает действовать стабилизирующий отбор, благоприятствующий сохранению оптимального генофонда, на базе которого формируются оптимально приспособленные к данным условиям фенотипы, которые становятся преобладающими в популяции. Процессы онтогенетического формообразования (морфогенезы), основывающиеся на разных генотипах, направляются стабилизирующим отбором в определенные русла, ведущие к формированию оптимального фенотипа (посредством распространения в популяции соответствующих генов-модификаторов). К.Уоддингтон назвал такое состояние канализованностью процессов морфогенеза.
В результате дикий тип, т.е. преобладающий в природной популяции оптимальный фенотип (или несколько фенотипов в полиморфной популяции), обычно объединяет генетически разнородных особей. Запас комбинативной изменчивости популяции благодаря канализованности процессов морфогенеза оказывается как бы скрытым за фенотипическим однообразием особей. Такое соотношение комбинативной и модификационной изменчивости оптимально при стабильных условиях среды, поскольку запас комбинативной изменчивости необходим для вида как "запас прочности", повышающий приспособляемость к изменениям среды обитания, а подгонка разных генотипов к одному или немногим оптимальным фенотипам повышает приспособленность к существующим в данный момент условиям. Состояние оптимальной стабильности популяции, которое обеспечивается ее специфическим генофондом и канализованностью процессов морфогенеза под контролем стабилизирующего отбора, носит название популяционного гомеостаза.
Стабилизирующий отбор направлен против крайних вариантов изменчивости, наблюдаемых в популяции, и благоприятствует консервативной части последней (рис. 12). В результате средняя величина (среднее состояние) признака остается из поколения в поколение неизменной. Стабилизирующий отбор ведет к большей фенотипической однородности популяции и к стабильности спектра ее изменчивости.

Очевидно, это направление отбора прямо противоположно схеме дивергентной эволюции, разработанной Ч.Дарвином. Под контролем стабилизирующего отбора, действующего длительное время, вид может внешне оставаться неизменным. Однако эта неизменность вида, в сущности, кажущаяся - она касается лини, постоянства оптимальных фенотипов. И.И.Шмальгаузен, разработавший концепцию стабилизирующего отбора, подчеркнул, что в таких условиях генофонд вида продолжает изменяться за счет накопления мутаций, фенотипическое проявление которых взаимно нейтрализуется (важную роль в этом играют гены-модификаторы), а также посредством накопления в гетерозиготном состоянии рецессивных мутаций. Таким образом, генофонд вида обогащается новыми аллелями, которые, оставаясь скрытыми, увеличивают мобилизационный резерв изменчивости, повышающий устойчивость вида к возможным изменениям внешних условий. Эти выводы подкрепляются результатами исследований наследственной изменчивости у разных видов, обитающих в стабильных внешних условиях - например у ряда морских животных (ракообразные, двустворчатые моллюски, иглокожие и другие группы). Оказалось, что генетическая гетерогенность популяций у видов, обитающих в стабильных условиях, не отличается от таковой у видов, обитающих в изменчивой среде.
При действии стабилизирующего отбора может происходить даже дивергенция видов, принимающая, однако, своеобразный характер. Механизм такого видообразования мы рассмотрим в следующей главе.
Направленный отбор наблюдается в том случае, если условия среды благоприятствуют какому-либо одному определенному направлению изменений признака, тогда как все другие варианты изменений подвергаются в той или иной степени негативному давлению отбора (рис. 13). В этом случае в популяции от поколения к поколению происходит сдвиг средней величины признака в определенном направлении. Результатом действия направленного отбора является постепенное изменение популяции в целом, происходящее посредством преобразования популяционного генофонда. В отличие от стабилизирующего отбора изменение частоты встречаемости различных аллелей и закрепление новых мутаций в генофонде при действии направленного отбора быстро отражаются на фенотипическом облике популяции. Это происходит тем быстрее, чем больше интенсивность отбора, если только последняя не превосходит приспособительных возможностей данной популяции - в последнем случае популяция вымирает.
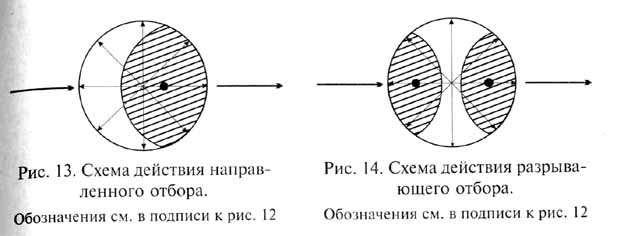
При направленном отборе особенно ярко проявляется воздействие отбора на темпы и преобладающее направление наследственной изменчивости (длящаяся изменчивость, по Ч.Дарвину): получают преимущества такие генотипы, которые обладают соответствующими возможностями дальнейших мутационных изменений, частота определенных мутаций может быть повышена с помощью генов-мутаторов; отбираются гены-модификаторы, способствующие доминантному фенотипическому проявлению благоприятных мутаций.
Осуществляемые под контролем направленного отбора эволюционные преобразования данной популяции затрагивают ее генофонд как целое; обособления каких-либо дочерних популяций (т.е. дивергенции) не происходит. Такую форму эволюционных преобразований вида Д.Симпсон назвал филетической эволюцией.
Разрывающий, или дизруптивный, отбор действует в том случае, когда условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака (рис. 14). В результате происходит возрастание изменчивости с последующим сдвигом среднего состояния признака в разных направлениях. Разрывающий отбор ведет либо к возникновению устойчивого адаптивного полиморфизма популяции, либо к ее дроблению, т. е. к разделению на несколько новых популяций такого же ранга. Последний вариант эволюционных изменений всего ближе к дарвиновской концепции дивергентной эволюции.

Тот или другой результат определяется конкретными условиями действия разрывающего отбора: при наличии какой-либо функциональной взаимозависимости между разными благоприятствуемыми генотипами в единой панмиктической популяции возникает полиморфизм, тогда как под влиянием различных вариантов условий в разных частях ареала при снижении скрещивания между населяющими их субпопуляциями происходит дивергенция последних. Д. Симпсон назвал последнюю форму эволюционных изменений видообразованием, поскольку она ведет к возникновению от одного родительского вида двух или нескольких новых - в противоположность филетической эволюции, преобразующей вид как целое, без разделения на дочерние виды.
При действии дизруптивного отбора у молодых дивергирую-щих видов соотношение фенотипической (модификационной) и комбинативной изменчивости становится противоположным тому состоянию, которое возникает под контролем стабилизирующего отбора: системы канализованности индивидуального развития разрушаются, в популяции происходит значительное усиление комбинативной и модификационной изменчивости, что приводит к возрастанию фенотипического разнообразия. Такая картина изменчивости была описана В.В.Кузнецовым в пределах комплекса видов сиговых рыб Coregonus muksun. Быстрое фе-нотипическое выявление разных вариантов комбинативной изменчивости повышает эффективность действия естественного отбора.
И разрывающий, и направленный отбор приводят к векторизованным изменениям генофонда и фенотипического облика популяций; сходно также их воздействие на онтогенез (разрушение систем его канализованности). Отдельные направления разрывающего отбора, действующие на разные аллопатрические популяции одного вида, можно рассматривать применительно к каждой данной популяции как направленный отбор. Поэтому иногда эти две формы отбора объединяют как движущий отбор, противопоставляемый стабилизирующему.
По отношению к разным адаптациям, различным признакам фенотипа у организмов одного вида отбор может иметь разное направление: для одних он оказывается движущим, для других -стабилизирующим. Когда давление движущего отбора по отношению к данной адаптации ослабевает, стабилизирующий отбор способствует закреплению достигнутых движущим отбором результатов, благоприятствуя оптимизации путей морфогенеза у новых адаптивных признаков для достижения максимальной надежности их воспроизведения в фенотипе (с повышением пенетрантно-сти и экспрессивности соответствующих аллелей посредством воздействия генов-модификаторов, а также механизма генокопиро-вания, см. гл. 1).
Одним из ярких примеров действия движущего отбора является развитие так называемого "индустриального меланизма", описанного более чем у 70 видов бабочек, распространенных в Англии и ряде других стран Европы, а также в США. Индустриальный меланизм представляет собой значительное повышение частоты встречаемости меланистических (имеющих темную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Особенно хорошо изучено это явление у березовой пяденицы Biston betularia (рис. 15) Англии.

До середины XIX в. в популяциях этого вида преобладали особи, имевшие светлую серовато-белую окраску с темными пятнышками, хорошо маскирующую бабочек на покрытых лишайниками стволах деревьев. В 1848 г. в Манчестере был отловлен первый экземпляр бабочки, имевший меланистическую окраску (морфа carbonaria). С тех пор частота встречаемости меланистических особей в популяциях этой бабочки, обитающих в промышленных районах Англии, постепенно возрастала, пока к началу XX в. количество меланистов не достигло в отдельных популяциях 95% всех особей. Популяции с преобладанием светлоокрашенных бабочек обитают ныне лишь на юго-западе Англии, в Северной Шотландии и в Ирландии. Варианты окраски у березовой пяденицы контролируются аллелями одного гена; аллель, обусловливающий темную окраску, имеет доминантное фенотипичес-кое проявление.
Возможно, меланистические особи в небольшой концентрации встречались в популяциях бабочек и до того момента, когда на них впервые обратили внимание ученые. Однако более широкому распространению аллеля, обусловливающего темную окраску насекомых, в те времена не благоприятствовали условия: ба-бочки-меланисты были гораздо лучше заметны на светлом фоне лишайников, покрывавших кору деревьев, чем "нормальные" светлоокрашенные особи. С развитием промышленности загрязнение воздуха вокруг промышленных центров привело к гибели лишайников на стволах деревьев, которые потемнели от копоти. В этих условиях меланистические формы бабочек оказались в лучших условиях, чем особи со светлой окраской, поскольку первые менее заметны на темных стволах деревьев, чем вторые. Это и определило быстрое увеличение концентрации бабочек-меланистов в популяциях промышленных районов. Благодаря меньшей "задымленности" воздуха в Ирландии, Северной Шотландии и Юго-Западной Англии там сохранилось прежнее преобладание светлой формы В. betularia.
Г. Кеттлуэлл в 50-60-х гг. XX в. экспериментально проверил эти соображения, выпустив определенное количество бабочек темной и светлой окраски в загрязненном лесу вблизи Бирмингема и затем повторив этот опыт в незагрязненном лесу в Дорсете. По данным Кеттлуэлла, птицы, питающиеся бабочками и, так сказать, непосредственно осуществляющие отбор последних, в Бирмингеме уничтожили значительно больше светлых бабочек, чем темных, тогда как в Дорсете наблюдалось обратное соотношение. Явление индустриального меланизма предоставляет ученым замечательные возможности для наблюдений за действием отбора в естественных популяциях. Этот эксперимент, "поставленный" природой при участии человека, уже показал, что изменения генофонда популяций могут происходить под контролем отбора чрезвычайно быстро - в течение нескольких десятилетий. Новые наблюдения показывают, что различия меланистических и светлых особей не сводятся только к разнице в окраске. Кеттлуэлл показал, что и те, и другие бабочки выбирают для отдыха субстраты предпочтительно неконтрастирующей окраски, т.е. ме-ланисты предпочитают темный, а "нормальные" особи - более светлый фон. Имеются также данные о различной жизнеспособности гусениц разных форм бабочек. Возможно, за сто с лишним лет, прошедшие со времени обнаружения первых бабочек-меланистов, не только перестроился генофонд отдельных популяций березовой пяденицы, но и произошли определенные преобразования внутри некоторых генных комплексов с участием генов-модификаторов, которые привели к повышению жизнеспособности и изменениям поведения у темных особей. Эти преобразования могли коснуться и степени доминантности фенотипического проявления того и другого аллелей в пользу повышения доминантности аллеля меланизма. Можно ожидать, что дальнейшее изучение индустриального меланизма и подобных ему явлений позволит проверить и многие другие положения теории отбора.
В обычном понимании отбор представляет собой преимущественное выживание и оставление потомства наиболее приспособленными особями. Можно сказать, что в этом аспекте отбор выступает как своего рода "эгоистический" фактор: видовые адаптации совершенствуются как адаптации отдельных особей. Однако, как мы уже подчеркивали в предшествующей главе, популяции и вид в целом являются целостными системами. Поэтому отбор должен также благоприятствовать закреплению и распространению признаков, которые сами по себе для отдельных особей не имеют приспособительной ценности или даже снижают их приспособленность, но обеспечивают существенные преимущества данной группировке особей как целому. Примеры действия такого "альтруистического" аспекта естественного отбора особенно многочисленны в поведении высших животных. Так возникли "сигналы тревоги", характерные для многих видов птиц: обнаружившая врага птица не затаивается в укрытии (что было бы наиболее благоприятно для ее собственной безопасности), а издает громкий крик, выдающий ее местонахождение хищнику, но предупреждающий других особей своего вида (часто сигнал тревоги воспринимается и другими видами). К этой же форме поведения принадлежит коллективная защита детенышей в стаях и стадах некоторых видов млекопитающих (слоны, копытные), защита и помощь другим особям своего вида (например, поддерживание на плаву раненых животных некоторыми китообразными).
При развитии подобных альтруистических особенностей поведения отбор оперирует не отдельными особями, а целыми их совокупностями (минимальной из них является так называемый дем - группа особей, непосредственно связанных в процессе размножения), благоприятствуя сохранению тех из них, в генофонде которых появляются "альтруистические" аллели. Этот аспект действия естественного отбора получил название "групповой отбор".
Концепция полового отбора была разработана Ч.Дарвином для объяснения развития у самцов некоторых видов животных украшающих признаков, нередко гипертрофированных, причудливых и ярких и поэтому отрицательно сказывающихся на защищенности этих особей и требующих для своего развития значительных энергетических затрат. Очевидно, предпочтение самками самцов с подобными признаками нуждается в объяснении. Первая попытка такого объяснения была сделана Р.Фишером, предположившим, что половой отбор, благоприятствующий развитию украшающих признаков, может действовать на основе предпочтения некоторыми самками таких признаков у самцов. В этом случае самцы, обладающие подобными украшениями, получают лучшие шансы оставить потомство, в котором самцы унаследуют от своих отцов эти признаки, а самки - от своих матерей соответствующие сексуальные предпочтения. Этот механизм (именуемый "фишеровскими процессами") обусловливает возникновение полового отбора, благоприятствующего дальнейшему развитию украшающих признаков у самцов. При этом первоначальное предпочтение самками определенных внешних признаков самцов может быть обусловлено особенностями физиологии их органов чувств (гипотеза первоначального сенсорного предпочтения).
А. Захави предложил интересное объяснение приспособительного значения украшающих признаков: степень их развития в целом должна соответствовать жизнеспособности самцов, поскольку такие "избыточные" структуры без ущерба для организма могут развивать лишь наиболее сильные индивиды с высоким уровнем обмена веществ и энергетических процессов, который контролируется генетически ("принцип гандикапа"). Тем самым степень развития у самцов таких украшений может служить для самок признаком, позволяющим выбрать наиболее жизнеспособных самцов - носителей "оптимальных" генов. Вероятно, развитие украшающих признаков под контролем полового отбора может происходить при совместном действии "фишеровских процессов" и "принципа гандикапа".
Факторы, влияющие на действие отбора
Действие того или иного типа (формы) отбора на каждый конкретный вид организмов определяется особенностями генетической системы данного вида, спецификой его взаимоотношений с внешней средой и характером изменений внешних условий. Очевидно, каждому из трех указанных выше основных направлений отбора благоприятствуют совершенно разные условия: постоянство условий существования - стабилизирующему отбору, изменение условий в одном направлении на всем ареале вида - направленному отбору, разнообразие условий существования (или изменение условий в разных направлениях) в пределах видового ареала - дизруптивному отбору. Вообще в филогенезе всякой группы организмов в течение ее длительной истории направление отбора многократно изменяется, хотя в некоторых случаях долгое время может преобладать какое-либо одно постоянное направление. Длительно действующий направленный отбор приводит к быстрым эволюционным преобразованиям значительного масштаба. Напротив, стабилизирующий отбор в некоторых случаях как бы "консервирует" фенотипический облик отдельных видов организмов в течение длительного времени.
В некоторых местообитаниях абиотические условия относительно мало изменялись или даже оставались в целом стабильными в течение многих миллионов лет (таковы, например, определенные зоны морского дна). В таких условиях некоторые виды организмов могут сохранять свой фенотипический облик практически неизменным в течение значительных периодов времени. Это так называемые персистирующие формы ("живые ископаемые"). Широко известными примерами таких организмов являются равноногие рачки-щитни Triops (рис. 16), организация которых не подвергалась заметным изменениям с триасового периода (более 200 млн лет); двустворчатые моллюски Leda, Nucula, Modiolus, существующие с каменноугольного периода (около 300 млн лет); плеченогие (брахиоподы) Lingula (рис. 17), не претерпевшие изменений с девонского периода(около 380 млн лет). Однако не следует рассматривать персистирующие виды как совершенно те же самые формы организмов,которые жили на Земле сотни миллионов лет назад: как мы уже отмечали, и под контролем стабилизирующего отбора генофонды видов продолжают изменяться, и с течением времени в них накапливается все больше мутаций структурных генов и соответствующи хмутаций генов-модификаторов, препятствующих изменениям фенотипа. Таким образом, в генетическом отношении современные персистирующие виды должны значительно отличаться от своих фЕнотипичЕски сходных предков.


Со времени Ч.Дарвина существенно изменились представления о факторах, благоприятствующих действию естественного отбора. Среди факторов, названных Дарвином (см. гл. 2, ч. I), универсальное значение имеет только длительность времени. Высокая изменчивость благоприятна в наибольшей степени для движущего отбора, стабилизирующий отбор действует против большого размаха изменчивости. Обширность ареала вида благоприятствует дизруптивному отбору.
Географическая изоляция территориальных (аллопатрических) популяций вида ныне рассматривается как один из важнейших факторов, способствующих действию разрывающего отбора. Некоторые ученые считают этот фактор непременным условием процесса видообразования. Под географической изоляцией понимается отделение данной популяции от других популяций родительского вида каким-либо географическим барьером, непроходимым для особей данного вида. Таким барьером может быть морской пролив, горный хребет, русло реки или даже просто район с непригодными для жизни данного вида условиями температуры, влажности, с отсутствием необходимой пищи и т. п.
Численность популяций и дрейф генов
Среди факторов, влияющих на эффективность отбора, особое место занимает численность популяций. С одной стороны, нельзя забывать, что действие отбора вообще имеет вероятностный характер (случайные причины могут привести к гибели или воспрепятствовать участию в размножении отдельных особей, являющихся носителями благоприятствуемых отбором признаков), поэтому для действия отбора необходима достаточно большая численность популяций. Однако, с другой стороны, в очень крупных панмиктических популяциях эффективность отбора может быть в определенной степени снижена благодаря относительной редкости выявления рецессивных мутаций в гомозиготном состоянии; обширный генофонд больших популяций более инертен, чем таковой популяций меньшего масштаба. Поэтому для действия движущего отбора наиболее благоприятны популяции средних размеров, но достаточно большие, чтобы отбор мог оказывать свое влияние вопреки действию случайных факторов.
Численность популяций в природе подвержена значительным колебаниям, зависящим от различных изменений внешней среды (таких, как засуха, морозная или бесснежная зима, урожай или неурожай кормовых растений, численность популяций других видов животных и т. п.). В урожайные годы численность популяций данного вида может существенно повыситься, вслед за чем при ухудшении условий наступает ее спад, тем более резкий, чем больше контраст условий в последовательные годы. Особенно ярко эти "популяционные волны" проявляются у видов с коротким жизненным циклом и быстрой сменой поколений (мелкие грызуны, ящерицы, лягушки, насекомые, однолетние растения, микроорганизмы и т.п.), но колебания численности популяций присущи всем без исключения видам организмов.
С.С.Четвериков подчеркнул большое эволюционное значение колебаний численности природных популяций, которые он назвал "волнами жизни". Поскольку для действия отбора важна численность популяций, очевидно, что ее изменения должны сопровождаться изменениями интенсивности, а в некоторых случаях - и направления отбора. Кроме того, колебания численности популяций неизбежно приводят к изменениям концентраций различных аллелей в популяционном генофонде, которые могут иметь случайный характер (исчезновение редких аллелей при снижении численности популяций или, наоборот, их значительное распространение в генофонде и т. п.). Такие изменения генофонда сами по себе представляют материал для действия отбора (отбор оптимального генофонда при последующей стабилизации условий).
В небольших популяциях, подвергшихся изоляции от других Популяций своего вида, влияние случайных факторов может выйти на первый план по отношению к действию отбора. Эта интересная ситуация в 1931 г. была описана Н.П.Дубининым и Д.Д.Ромашовым под названием "генетико-автоматические процессы" и, независимо от них, в том же году С. Райтом, который обозначил ее термином "дрейф генов". Если небольшая популяция (менее 50 особей) оказывается изолированной, прежде всего проявляется эффект, названный "принципом основателя". Генофонд малочисленной популяции с самого начала беднее, чем генофонд родительского вида, и подбор аллелей в генофонде "популяции основателей" случаен из-за случайного подбора особей. Такой генофонд не сбалансирован, поскольку он возник на основе случайности, а не под контролем естественного отбора. Так как отбор действует на вероятностном уровне и требует для своей эффективности достаточной численности особей, дальнейшая судьба генофонда маленькой популяции определяется в первую очередь действием различного рода случайных факторов.
Общей тенденцией будет выпадение из генофонда малой популяции наиболее редких аллелей, сопровождающееся их замещением аллелями, которые с самого начала были более многочисленными. Это ведет к дальнейшему обеднению генофонда малой популяции и в результате к его гомозиготизации. Важно подчеркнуть, что указанные изменения генофонда не имеют приспособительного характера: утраченные аллели могут обладать равной и даже большей приспособительной ценностью, чем аллели, ставшие в популяции преобладающими: ведь потеря первых определяется не действием отбора, а случайной гибелью особей. Такая случайная гибель особей от самых различных причин (хищники, заболевания, факторы абиотической среды) происходит всегда и в любой популяции, но при большом количестве особей она отступает на задний план - в среднем успешнее выживают и оставляют потомство лучше приспособленные организмы. В малой же популяции фактор случайности становится основным. Естественно, что в этом случае лучшие шансы для сохранения имеют первоначально более многочисленные аллели вне зависимости от их приспособительной ценности.
Таким образом, действие случайных факторов обедняет и значительно изменяет генофонд малой изолированной популяции по сравнению с его исходным состоянием. Это явление и называется собственно генетико-автоматическими процессами или дрейфом генов. В результате подобных процессов может сложиться жизнеспособная популяция со своеобразным генофондом (случайно, поскольку отбор в данном случае не играл ведущей роли!). Спектр изменчивости этой популяции существенно отличается от такового родительского вида. С постепенным увеличением численности особей вновь восстановится эффективность естественного отбора, который будет действовать уже на новый генофонд, обладающий специфическим спектром изменчивости. Совокупность всех этих процессов может привести к обособлению нового вида.
Соотношения роли отбора и дрейфа генов при разной численности популяций можно выразить по С.Райту следующим образом:
если численность популяции N<1/(2S) (где S- коэффициент интенсивности отбора, см. выше), преобладает воздействие дрейфа генов; если N>1/(4S) преобладает отбор; в диапазоне N от 1/(4S) до 1/(2S) совместно действуют оба фактора. Так, при умеренной интенсивности отбора (S=0,01) изменения соотношений любой пары аллелей регулируются дрейфом генов при /V<50; при слабой интенсивности отбора (S= 0,001) дрейф генов оказывается определяющим фактором при N< 500 и т.д.
В соответствии с этим генетико-автоматические процессы могут существенно влиять на генофонд популяции основателей в первое время после ее закрепления в новом районе, пока она еще достаточно малочисленна; затем на первый план неизбежно выходит действие естественного отбора. Если условия в новом районе останутся благоприятными, численность популяции быстро увеличится и эффект случайности - дрейф генов - будет все более затушевываться влиянием отбора, как бы ни было слабо его давление. Если же условия среды станут неблагоприятными и численность популяции вследствие этого останется небольшой - это явится непосредственным результатом жесткого давления отбора и дрейф опять-таки не сможет быть определяющим фактором эволюционных перестроек генофонда в течение длительного времени.
Сравнивая эволюционное значение рассмотренных нами факторов, мы можем заключить, что наличие мутаций и естественного отбора необходимо и достаточно, чтобы обеспечить приспособительную и дивергентную эволюцию организмов. Поэтому мутационный процесс и естественный отбор можно обозначить как необходимые факторы эволюционного процесса. При этом отбор является единственным известным ныне фактором эволюции, который объединяет движущее, направляющее и интегрирующее воздействия на организмы, формируя их приспособления и влияя на саму мутационную изменчивость. Прочие рассмотренные нами элементарные эволюционные факторы (колебания численности, обмен генетической информацией между разными популяциями, географическая изоляция, дрейф генов) являются дополнительными по отношению к мутационному процессу и естественному отбору. Разумеется, для полного понимания эволюционного процесса необходимо учитывать сложные взаимодействия всех указанных элементарных эволюционных факторов.
Дата добавления: 2015-04-16; просмотров: 353; Мы поможем в написании вашей работы!; Нарушение авторских прав |