КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ОБЪЯСНЯЮЩИЕ МЕХАНИЗМЫ СТАРЕНИЯ
Геронтология знает не менее 500 гипотез, объясняющих и первопричину, и механизмы старения организма. Подавляющее большинство их не выдержало проверки временем и представляет чисто исторический интерес. К ним, в частности, относятся гипотезы, связывающие старение с расходованием особого вещества клеточных ядер, страхом смерти, утратой некоторых невосполняемых веществ, получаемых организмом в момент оплодотворения, самоотравлением продуктами жизнедеятельности, токсичностью продуктов, образуемых под действием микрофлоры толстого кишечника. Гипотезы, представляющие научную ценность в наши дни, соответствуют одному из двух главных направлений.
Некоторые авторы рассматривают старение как стохастический процесс возрастного накопления «ошибок», неизбежно случающихся в ходе обычных процессов жизнедеятельности, а также повреждений биологических механизмов под действием внутренних (спонтанные мутации) или внешних (ионизирующее облучение) факторов. Стохастичность обусловливается случайным характером изменений во времени и локализации в организме. В различных вариантах гипотез данного направления первостепенная роль отводится разным внутриклеточным структурам, от первичного повреждения которых зависят функциональные расстройства на клеточном, тканевом и органном уровнях. Прежде всего это генетический аппарат клеток (гипотеза соматических мутаций).
Многие исследователи связывают начальные изменения старения организма с изменениями строения и, следовательно, физико-химических и биологических свойств макромолекул: ДНК, РНК, белков хроматина, цитоплазматических и ядерных белков, ферментов. Особо выделяют также липиды клеточных мембран, часто являющиеся мишенью для свободных радикалов. Сбои в работе рецепторов, в частности клеточных оболочек, нарушают эффективность регуляторных механизмов, что приводит к рассогласованию процессов жизнедеятельности.
К рассматриваемому направлению относятся также гипотезы, усматривающие первооснову старения в нарастающем с возрастом износе структур в диапазоне от макромолекул до организма в целом, приводящем в конце концов к состоянию, не совместимому с жизнью. Такое представление, однако, слишком прямолинейно. Напомним, что возникновению и накоплению мутационных изменений в ДНК противостоят природные антимутационные механизмы, а вредные последствия образования свободных радикалов снижаются благодаря функционированию антиоксидантных механизмов. Таким образом, если «концепция износа» биологических структур правильно отражает сущность старения, то итог в виде большей или меньшей скорости старческих изменений возраста, в котором у разных людей эти изменения становятся очевидными, является следствием наложения разрушительных и защитных процессов. В этом случае гипотеза износа с неизбежностью включает в себя такие факторы, как генетическая предрасположенность, условия и даже образ жизни, от которых, как мы видели, зависит скорость старения.
Второе направление представлено генетическими или программными гипотезами, согласно которым процесс старения находится под прямым генетическим контролем. Указанный контроль, согласно одним взглядам, осуществляется с помощью специальных генов. По другим взглядам, он связан с наличием специальных генетических программ, как это имеет место в отношении других стадий онтогенеза, например эмбриональной.
В пользу запрограммированности старения приводят доказательства, многие из которых уже рассмотрены в разд. 8.6.1. Обычно также ссылаются на наличие в природе видов, у которых вслед за размножением бурно нарастают изменения, приводящие животных к гибели. Типичный пример —тихоокеанские лососи (нерка, горбуша), погибающие после нереста. Пусковой механизм в этом случае связан с изменением режима секреции половых гормонов, что следует рассматривать как особенность генетической программы индивидуального развития лососевых, отражающей их экологию, а не как универсальный механизм старения. Примечательно, что кастрированная горбуша не нерестится и живет в 2—3 раза дольше. Именно в эти дополнительные годы жизни следует ожидать появления признаков старения в клетках и тканях.
Некоторые программные гипотезы основаны на допущении, что в организме функционируют биологические часы, в соответствии с которыми происходят возрастные изменения. Роль «часов» приписывают, в частности, вилочковой железе, прекращающей функционирование при переходе организма в зрелый возраст. Еще один кандидат — это нервная система, особенно некоторые ее отделы (гипоталамус, симпатическая нервная система), главным функциональным элементом которой являются первично стареющие нервные клетки. Допустим, что прекращение в определенном возрасте функций тимуса, что, несомненно, находится под генетическим контролем, является сигналом начала старения организма. Это, однако, не означает генетического контроля процесса старения. В отсутствие тимуса ослабляется иммунологический контроль за аутоиммунными процессами. Но для того чтобы эти процессы пошли, необходимы либо мутантные лимфоциты (повреждения ДНК), либо белки с измененной структурой и антигенными свойствами.
Генетические программы, в том числе и индивидуального развития, являются всегда результатом эволюции, закрепляемым в генофонде вида вследствие естественного отбора. На первый взгляд, естественный отбор должен благоприятствовать увеличению продолжительности жизни. В связи с этим приобретение видом в ходе эволюции генетической программы старения, обусловливающего неизбежность смерти, представляется маловероятным. Рассмотрим следующий пример. В природных условиях в первый год жизни сохраняется в живых лишь 1/4 синиц каждого поколения. По истечении 2-го года от поколения остаются единицы, если это вообще происходит. В лабораторных условиях птицы достигают 9-летнего возраста. В таком случае практически невозможно объяснить, в силу каких обстоятельств естественный отбор мог формировать генетическую программу саморазрушения организма в процессе старения, рассчитанную на 7—8 лет жизни, которые синицами не проживаются.
Изложенное выше не исключает зависимости скорости старения и времени наступления старческих изменений от генетических факторов, однако этими факторами не являются специальные гены или программа. Рассмотрим еще один пример. Для хореи Гентингтона типичным признаком служит сильный тремор (дрожание) головы, конечностей (пляска святого Витта). Симптомы этого наследственного заболевания обычно появляются в возрасте 35—39 лет, причем у мужчин позднее, чем у женщин. Различие в сроках появления болезни объясняется особенностями эволюции мужского и женского генотипов. У мужчин, имеющих по сравнению с женщинами большую продолжительность репродуктивного периода, давление отбора против соответствующего признака угасает с возрастом более медленно. Неблагоприятное фенотипическое действие гена, лежащего в основе хореи Геттингтона, в юношеском и зрелом возрасте подавлялось благодаря присутствию в геноме генов-модификаторов (см. разд. 8.6.1).
Таким образом, из двух принципиально различных направлений в объяснении старения как закономерной стадии онтогенеза в настоящее время более обоснованным является представление, рассматривающее этот процесс как износ биологических структур, а не генетически предопределенное саморазрушение.
67.Основные концепции в биологии развития (преформизм, эпигенез).
Вопрос об индивидуальном развитии — онтогенезе — привлекал к себе внимание со времен Аристотеля. Благодаря усилиям многих исследователей к XVII в. был накоплен обширный материал по изменениям, происходящим с зародышами позвоночных на макроуровне. Появление в XVII столетии [микроскопа] перевело эмбриологию на качественно новый уровень, хотя несовершенство первых микроскопов и крайне примитивная техника изготовления микропрепаратов делали практически недоступными для изучения ранние стадии развития зародышей.
В XVII-XVIII вв. оформились два взгляда на онтогенез — преформизм и эпигенез. Сторонники преформизма полагали, что зародышевое развитие сводится к росту вполне сформированного зародыша. Предполагалось, что зародыш — уменьшенный вариант сложноустроенного взрослого организма — существовал в такой форме с момента творения. Преформисты, в свою очередь, разделились на две группы. Овисты — Я. Сваммердам, А. Валлиснери, М. Мальпиги, Ш. Бонне, А. Галлер, Л. Спаланзани и др. — считали, что уже сформированный зародыш находится в яйцеклетке, а мужское половое начало лишь дает толчок к развитию. Анималькулисты (А. Левенгук, Н. Хартсекер, И.Н. Либеркюн и др.) утверждали, что зародыш заключен в сперматозоиде, который развивается за счет питательных веществ яйца. А. Левенгук допускал существование "мужских" и "женских" сперматозоидов. Крайним выражением преформизма явилась теория вложений. Согласно ей, половые клетки зародышей, как матрешки, уже несут в себе зародышей следующего поколения, в тех содержатся зародыши последующих поколений, и так далее.
Воззрения преформистов базировались на некоторых фактических данных. Так, Я. Сваммердам, вскрыв куколку бабочки, обнаружил там вполне сформированное насекомое. Ученый воспринял это как доказательство того, что более поздние стадии развития "спрятаны" в более ранних и до поры до времени не видны. Ш. Бонне открыл партеногенез — размножение без оплодотворения — у тлей, что было воспринято овистами как довод в их пользу.
Ряд фактов с точки зрения преформизма получал не слишком удачное объяснение. В XVIII в. благодаря работам Р. Реомюра и А. Трамбле становится широко известным явление регенерации. Ш. Бонне сводил и регенерацию, и различные формы бесполого размножения к "предсуществованию" в организме особых, специально на сей предмет предобразованных зародышей. Таким образом, гидра вся целиком с точки зрения преформизма представляла собой яичник или собрание зародышей. Сходство детей с обоими родителями преформисты объясняли тем, что зародыш, происшедший из яйца или из семенного анималькуля, формируется по образу и подобию своих родителей под влиянием воображения матери в течение утробной жизни. Плохо согласовывалось с идеей преформизма и появление уродств. Впрочем, некоторые сторонники этой концепции допускали, что вложенные зародыши не обязательно идентичны друг с другом, вплоть до того, что и сам прогресс живых форм мог быть преформирован в момент творения.
Приверженцы альтернативного течения — эпигенетики — считали, что в процессе онтогенеза происходит новообразование структур и органов зародыша из бесструктурного вещества. Впервые идея эпигенеза встречается в труде В. Гарвея "Исследования о нарождении животных" (1651 г.), но в полной мере соответствующие взгляды были выражены К.Ф. Вольфом (1733-1794). К.Ф. Вольф исходил из того, что, если правы преформисты, то все органы зародыша, как только мы их можем увидеть, должны быть полностью сформированы. В своей работе "Теория зарождения" (1759 г.) ученый описывает картины постепенного возникновения различных органов из "неорганизованной массы" у животных и растений. К сожалению, К.Ф. Вольф работал с довольно плохим микроскопом, что породило многие фактические неточности, но это не умаляет значения созданной им теории эпигенеза.
Эпигенетической точки зрения в XVIII в. придерживались П. Мопертюи, Дж. Нидхэм, Д. Дидро, отчасти Ж. Бюффон.
Решающий перелом в споре между представителями двух течений произошел в XIX в. после работ К.М. Бэра (1792-1876), сумевшего снять альтернативу — или преформизм, или эпигенез. К.М. Бэр считал, что нигде в зародыше не происходит новообразований, имеют место лишь преобразования. При этом преобразование К.М. Бэр понимал отнюдь не в духе преформизма, а рассматривал его как подлинное развитие, с глубокими качественными преобразованиями от более простого и недифференцированного к более сложному и дифференцированному.
68.Механизмы онтогенеза на клеточном уровне: пролиферация, миграция, адгезия, избирательная сортировка клеток, дифференцировка, запрограммированная гибель клеток).
69.Взаимодействие частей развивающегося организма. Эмбриональная индукция, её виды.
70.Целостность онтогенеза. Эмбриональная регуляция. Детерминация частей развивающегося организма. Морфогенез.
71.Роль наследственности и среды в онтогенезе. Критические периоды в онтогенезе человека. Тератогенез, канцерогенез. Аномалии и пороки развития.
72.Пороки развития в пренатальном периоде онтогенеза человека. Классификация пороков развития. Наследственные и ненаследственные пороки. Фенокопии.
73.Регенерация. Физиологическая регенерация, её значение.
Регенерация – процесс вторичного развития органа или ткани, вызванный повреждениями какого – либо рода.
Физиологическая регенерация – восстановление частей организма, износившихся в процессе жизнедеятельности. Действует на протяжении всего онтогенеза, поддерживает постоянство структур, несмотря на гибель клеток. Интенсивные процессы физиологической регенерации при восстановлении клеток крови, эпидермиса, слизистых оболочек. Примерами могут быть линька птиц, рост зубов у грызунов. Физиологическая регенерация происходит не только в тканях с интенсивно делящимися клетками, но и там, где клетки делятся незначительно. 25 гепатоцитов из 1000 погибают и столько же восстанавливаются. Физиологическая регенерация – динамический процесс, который включает в себя клеточное деление и другие процессы. Обеспечение функций лежит в основе нормального функционирования организма.
Физиологическая регенерация представляет собой процесс обновления функционирующих структур организма. Благодаря физиологической регенерации поддерживается структурный гомеостаз и обеспечивается возможность постоянного выполнения органами их функций. С общебиологической точки зрения, физиологическая регенерация, как и обмен веществ, является проявлением такого важнейшего свойства жизни, как самообновление.
Примером физиологической регенерации на внутриклеточном уровне являются процессы восстановления субклеточных структур в клетках всех тканей и органов. Значение ее особенно велико для так называемых «вечных» тканей, утративших способность к регенерации путем деления клеток. В первую очередь это относится к нервной ткани.
Примерами физиологической регенерации на клеточном и тканевом уровнях являются обновление эпидермиса кожи, роговицы глаза, эпителия слизистой кишечника, клеток периферической крови и др. Обновляются производные эпидермиса — волосы и ногти. Это так называемая пролиферативная регенерация, т.е. восполнение численности клеток за счет их деления. Во многих тканях существуют специальные камбиальные клетки и очаги их пролиферации. Это крипты в эпителии тонкой кишки, костный мозг, пролиферативные зоны в эпителии кожи. Интенсивность клеточного обновления в перечисленных тканях очень велика. Это так называемые «лабильные» ткани. Все эритроциты теплокровных животных, например, сменяются за 2—4 мес, а эпителий тонкой кишки полностью сменяется за 2 сут. Это время требуется для перемещения клетки из крипты на ворсинку, выполнения ею функции и гибели. Клетки таких органов, как печень, почка, надпочечник и др., обновляются значительно медленнее. Это так называемые «стабильные» ткани.
Об интенсивности пролиферации судят по количеству митозов, приходящихся на 1000 подсчитанных клеток. Если учесть, что сам митоз в среднем длится около 1 ч, а весь митотаческий цикл в соматических клетках в среднем протекает 22—24 ч, то становится ясно, что для определения интенсивности обновления клеточного состава тканей необходимо подсчитать количество митозов в течение одних или нескольких суток. Оказалось, что количество делящихся клеток не одинаково в разные часы суток. Так был открыт суточный ритм клеточных делений.
Суточный ритм количества митозов обнаружен не только в нормальных, но и в опухолевых тканях. Он является отражением более общей закономерности, а именно ритмичности всех функций организма. Одна из современных областей биологии — хронобиология — изучает, в частности, механизмы регуляции суточных ритмов митотической активности, что имеет весьма важное значение для медицины. Существование самой суточной периодичности количества митозов указывает на регулируемость физиологической регенерации организмом. Кроме суточных существуют лунные и годичные циклы обновления тканей и органов.
В физиологической регенерации выделяют две фазы: разрушительную и восстановительную. Полагают, что продукты распада части клеток стимулируют пролиферацию других. Большую роль в регуляции клеточного обновления играют гормоны.
Физиологическая регенерация присуща организмам всех видов, но особенно интенсивно она протекает у теплокровных позвоночных, так как у них вообще очень высока интенсивность функционирования всех органов по сравнению с другими животными.
74.Репаративная регенерация, её значение. Способы репаративной регенерации. Проявление регенерационной способности в филогенезе. Молекулярно-генетические, клеточные и системные механизмы регенерации. Особенности восстановительных процессов у млекопитающих.
Регенерация может быть физиологической и репаративной. В свою очередь репаративная регенерация бывает нескольких видов:
- возмещающая;
- посттравматическая;
- восстановительная;
- патологическая.
Репаративная регенерация – восстановление поврежденных тканей и органов после чрезвычайных воздействий. При полной регенерации восстанавливается полное исходное строение ткани после ее повреждения, её архитектура остается неизменной. Распространена у организмов, способных к бесполому размножению. Например, белая планария, гидра, моллюски (если удалить голову, но оставить нервно – узловую структуру). Типичная репаративная регенерация возможна у высших организмов, в т.ч. и человека. Например, при устранении некротических клеток органов. В острой стадии пневмонии происходит деструкция альвеол и бронхов, затем происходит восстановление. При действии гепатотропных ядов возникают диффузные некротические изменения печени. После прекращения действия ядов восстанавливается архитектоника за счет деления гепатоцитов – клеток печеночной паренхимы. Восстанавливается исходная структура. Гомоморфоз – восстановление структуры в том виде, в котором она существовала до разрушения. Неполная репаративная регенерация – регенерированный орган отличается от удаленного - гетероморфоз. Исходная структура не восстанавливается, а иногда вместо одного органа развивается другой орган. Например, глаз у рака. При удалении в некоторых случаях развивается антенна. У человека печень при удалении части печеночной доли аналогично регенерирует. Возникает рубец и через 2 - 3 месяца после операции масса печени восстанавливается, а восстановления формы органа не происходит. Это происходит из-за удаления и повреждения соединительной ткани во время операции.
У млекопитающих могут регенерировать все 4 вида ткани.
1. Соединительная ткань. Рыхлая соединительная ткань обладает высокой способностью к регенерации. Лучше всего регенерируют интерстициальные компоненты – образуется рубец, замещающийся тканью. Костная ткань – аналогично. Основные элементы, восстанавливающие ткань – остеобласты (малодифференцированные камбиальные клетки костной ткани);
2. Эпителиальная ткань. Обладает выраженной регенерационной реакцией. Эпителий кожи, роговая оболочка глаза, слизистые оболочки полости рта, губ, носа, желудочно-кишечного тракта, мочевого пузыря, слюнные железы, паренхима почек. При наличии раздражающих факторов могут происходить патологические процессы, приводящие к разрастанию тканей, что приводит к раковым опухолям.
3. Мышечная ткань. Значительно меньше регенерирует, чем эпителиальная и соединительная ткани. Поперечная мускулатура – амитоз, гладкая – митоз. Регенерирует за счет недифференцированных клеток – сателлитов. Могут разрастаться и регенерировать отдельные волокна, и даже целые мышцы.
4. Нервная ткань. Обладает плохой способностью к регенерации. В эксперименте показано, что клетки периферической и вегетативной нервной системы, двигательные и чувствительные нейроны в спинном мозге мало регенерируют. Аксоны хорошо регенерируют за счет Шванновских клеток. В головном мозге вместо них - глия, поэтому регенерация не происходит.
При регенерации миокарда и центральной нервной системы сначала образуется рубец, а затем идет регенерация за счет увеличения размеров клеток, внутриклеточная регенерация также имеет место. Клетки миокарда митозом не делятся. Разница происходит из-за развития в эмбриональном периоде. У взрослых организмов очень мощно функционирует ЭПР и это тормозит клеточное деление.
Процесс регенерации конечности у тритона/ саламандры.
После ампутации регенерация конечности происходит строго упорядоченно, всегда одинаково. Восстанавливающийся конец округляется, затем приобретает коническую форму, растет в длину, становится похожим на ласт. Потом закладываются пальцы. К 8 неделе регенерация конечности полностью завершена.
На клеточном уровне выделяют несколько фаз регенерации конечности:
1) фаза заживления раны;
2) процесс демонтирования;
3) фаза « конической бластемы»;
4) фаза редифференцировки.
Фаза заживления раны. В этот период происходит обрастание клетками раны на культе, возникает апикальная «шапочка» (если контакт нарушен – регенерации не будет).
Процесс демонтирования. После заживления, в тканях, прилежащих к культе, происходит рассасывание ткани. Мышечные волокна утрачивают упорядоченность, становятся «растрепанными». В костной ткани утрачивается надкостница, появляются гигантские фагоцитирующие клетки, имеющие не менее 3-х ядер. Эти клетки захватывают матрикс и освобождают место для роста новой кости и хряща, удаляя ненужный материал. Концевая часть культи становится отечной и выпячивается. В культе накапливаются однотипные дедифференцированные клетки, уподобленные эмбриональным клеткам. Через некоторое время начинается деление дедифференцированных клеток.
В отрастающую культю врастают нервы, и наступает стадия « конической бластемы». Конечность имеет форму ласта, нарастает клеточная масса, восстанавливается кровоток. Возникает «регенерационная почка».
Фаза редифференцировки. Конечность удлиняется, начинается редифференцировка, и процесс регенерации подходит к концу. Если денервировать конечность - регенерация не произойдет т.к. нервная ткань выполняет эндокринную, проводящую функции. Кроме того, нервная ткань осуществляет секрецию белкового гормона, под контролем которого осуществляется регенерация.
Репаративная (от лат. reparatio — восстановление) регенерация наступает после повреждения ткани или органа. Она очень разнообразна по факторам, вызывающим повреждения, по объемам повреждения, по способам восстановления. Механическая травма, например оперативное вмешательство, действие ядовитых веществ, ожоги, обморожения, лучевые воздействия, голодание, другие болезнетворные агенты,— все это повреждающие факторы. Наиболее широко изучена регенерация после механической травмы. Способность некоторых животных, таких, как гидра, планария, некоторые кольчатые черви, морские звезды, асцидия и др., восстанавливать утраченные органы и части организма издавна изумляла ученых. Ч. Дарвин, например, считал удивительными способность улитки воспроизводить голову и способность саламандры восстанавливать глаза, хвост и ноги именно в тех местах, где они отрезаны.
Объем повреждения и последующее восстановление бывают весьма различными. Крайним вариантом является восстановление целого организма из отдельной малой его части, фактически из группы соматических клеток. Среди животных такое восстановление возможно у губок и кишечнополостных. Среди растений возможно развитие целого нового растения даже из одной соматической клетки, как это получено на примере моркови и табака. Такой вид восстановительных процессов сопровождается возникновением новой морфогенетической оси организма и назван Б.П. Токиным «соматическим эмбриогенезом», ибо во многом напоминает эмбриональное развитие.
Существуют примеры восстановления больших участков организма, состоящих из комплекса органов. В качестве примера служат регенерация ротового конца у гидры, головного конца у кольчатого червя и восстановление морской звезды из одного луча (рис. 8.24). Широко распространена регенерация отдельных органов, например конечности у тритона, хвоста у ящерицы, глаз у членистоногих. Заживление кожных покровов, ран, повреждений костей и других внутренних органов является менее объемным процессом, но не менее важным для восстановления структурно-функциональной целостности организма. Особый интерес представляет способность зародышей на ранних стадиях развития восстанавливаться после значительной утраты материала. Эта способность была последним аргументом в борьбе между сторонниками преформизма и эпигенеза и привела в 1908 г. Г. Дриша к концепции эмбриональной регуляции.

Рис. 8.24. Регенерация комплекса органов у некоторых видов беспозвоночных животных. А — гидра;Б — кольчатый червь; В — морская звезда
(пояснение см. в тексте)
Существует несколько разновидностей или способов репаративной регенерации. К ним относят эпиморфоз, морфаллаксис, заживление эпителиальных ран, регенерационную гипертрофию, компенсаторную гипертрофию.
Эпителизация при заживлении ран с нарушенным эпителиальным покровом идет примерно одинаково, независимо от того, будет далее происходить регенерация органа путем эпиморфоза или нет. Эпидермальное заживление раны у млекопитающих в том случае, когда раневая поверхность высыхает с образованием корки, проходит следующим образом (рис. 8.25). Эпителий на краю раны утолщается вследствие увеличения объема клеток и расширения межклеточных пространств. Сгусток фибрина играет роль субстрата для миграции эпидермиса в глубь раны. В мигрирующих эпителиальных клетках нет митозов, однако они обладают фагоцитарной активностью. Клетки с противоположных краев вступают в контакт. Затем наступает кератинизация раневого эпидермиса и отделение корки, покрывающей рану.

Рис. 8.25. Схема некоторых событий, происходящих
при эпителизации кожной раны у млекопитающих.
А—начало врастания эпидермиса под некротическую ткань; Б— срастание эпидермиса и отделение струпа:
1—соединительная ткань, 2—эпидермис, 3—струп, 4—некротическая ткань
К моменту встречи эпидермиса противоположных краев в клетках, расположенных непосредственно вокруг края раны, наблюдается вспышка митозов, которая затем постепенно падает. По одной из версий, эта вспышка вызвана понижением концентрации ингибитора митозов — кейлона.
Эпиморфоз представляет собой наиболее очевидный способ регенерации, заключающийся в отрастании нового органа от ампутационной поверхности. Регенерация конечности тритона и аксолотля изучена детально. Выделяют регрессивную и прогрессивную фазы регенерации. Регрессивная фаза начинается с заживления раны, во время которого происходят следующие основные события: остановка кровотечения, сокращение мягких тканей культи конечности, образование над раневой поверхностью сгустка фибрина и миграция эпидермиса, покрывающего ампутационную поверхность.
Затем начинается разрушение остеоцитов на дистальном конце кости и других клеток. Одновременно в разрушенные мягкие ткани проникают клетки, участвующие в воспалительном процессе, наблюдается фагоцитоз и местный отек. Затем вместо образования плотного сплетения волокон соединительной ткани, как это происходит при заживлении ран у млекопитающих, в области под раневым эпидермисом утрачиваются дифференцированные ткани. Характерна остеокластическая эрозия кости, что является гистологическим признаком дедифференцировки. Раневой эпидермис, уже пронизанный регенерирующими нервными волокнами, начинает быстро утолщаться. Промежутки между тканями все более заполняются мезенхимоподобными клетками. Скопление мезенхимных клеток под раневым эпидермисом является главным показателем формирования регенерационной бластемы. Клетки бластемы выглядят одинаково, но именно в этот момент закладываются основные черты регенерирующей конечности.
Затем начинается прогрессивная фаза, для которой наиболее характерны процессы роста и морфогенеза. Длина и масса регенерационной бластемы быстро увеличиваются. Рост бластемы происходит на фоне идущего полным ходом формирования черт конечности, т.е. ее морфогенеза. Когда форма конечности в общих чертах уже сложилась, регенерат все еще меньше нормальной конечности. Чем крупнее животное, тем больше эта разница в размерах. Для завершения морфогенеза требуется время, по истечении которого регенерат достигает размеров нормальной конечности.
Некоторые стадии регенерации передней конечности у тритона после ампутации на уровне плеча показаны на рис. 8.26. Время, необходимое для полной регенерации конечности, варьирует в зависимости от размера и возраста животного, а также от температуры, при которой она протекает.

Рис. 8.26. Стадии регенерации передней конечности у тритона
У молодых личинок аксолотлей конечность может регенерировать за 3 нед, у взрослых тритонов и аксолотлей за 1—2 мес, а у наземных амбистом для этого требуется около 1 года.
При эпиморфной регенерации не всегда образуется точная копия удаленной структуры. Такую регенерацию называют атипичной. Существует много разновидностей атипичной регенерации. Гипоморфоз — регенерация с частичным замещением ампутированной структуры. Так, у взрослой шпорцевой лягушки возникает шиловидная структура вместо конечности. Гетероморфоз — появление иной структуры на месте утраченной. Это может проявляться в виде гомеозисной регенерации, заключающейся в появлении конечности на месте антенн или глаза у членистоногих, а также в изменении полярности структуры. Из короткого фрагмента планарии можно стабильно получать биполярную планарию (рис. 8.27).
Встречается образование дополнительных структур, или избыточная регенерация. После надреза культи при ампутации головного отдела планарии возникает регенерация двух голов или более (рис. 8.28). Можно получить больше пальцев при регенерации конечности аксолотля, повернув конец культи конечности на 180°. Дополнительные структуры являются зеркальным отражением исходных или регенерировавших структур, рядом с которыми они расположены (закон Бэйтсона).
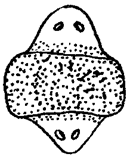
Рис. 8.27. Биполярная планария
Морфаллаксис — это регенерация путем перестройки регенерирующего участка. Примером служит регенерация гидры из кольца, вырезанного из середины ее тела, или восстановление планарии из одной десятой или двадцатой ее части. На раневой поверхности в этом случае не происходит значительных формообразовательных процессов. Отрезанный кусочек сжимается, клетки внутри него перестраиваются, и возникает целая особь уменьшенных размеров, которая затем растет. Этот способ регенерации впервые описал Т. Морган в 1900 г. В соответствии с его описанием морфаллаксис осуществляется без митозов. Нередко имеет место сочетание эпиморфного роста на месте ампутации с реорганизацией путем морфаллаксиса в прилежащих частях тела.

Рис. 8.28. Многоголовая планария, полученная после ампутации головы
и нанесения насечек на культю
Регенерационная гипертрофия относится к внутренним органам. Этот способ регенерации заключается в увеличении размеров остатка органа без восстановления исходной формы. Иллюстрацией служит регенерация печени позвоночных, в том числе млекопитающих. При краевом ранении печени удаленная часть органа никогда не восстанавливается. Раневая поверхность заживает. В то же время внутри оставшейся части усиливается размножение клеток (гиперплазия) и в течение двух недель после удаления 2/3 печени восстанавливаются исходные масса и объем, но не форма. Внутренняя структура печени оказывается нормальной, дольки имеют типичную для них величину. Функция печени также возвращается к норме.
Компенсаторная гипертрофия заключается в изменениях в одном из органов при нарушении в другом, относящемся к той же системе органов. Примером является гипертрофия в одной из почек при удалении другой или увеличение лимфатических узлов при удалении селезенки.
Последние два способа отличаются местом регенерации, но механизмы их одинаковы: гиперплазия и гипертрофия.
Восстановление отдельных мезодермальных тканей, таких, как мышечная и скелетная, называют тканевой регенерацией. Для регенерации мышцы важно сохранение хотя бы небольших ее культей на обоих концах, а для регенерации кости необходима надкостница. Регенерация путем индукции происходит в определенных мезодермальных тканях млекопитающих в ответ на действие специфических индукторов, которые вводят внутрь поврежденной области. Этим способом удается получить полное замещение дефекта костей черепа после введения в него костных опилок.
Таким образом, существует множество различных способов или типов морфогенетических явлений при восстановлении утраченных и поврежденных частей организма. Различия между ними не всегда очевидны, и требуется более глубокое понимание этих процессов.
Изучение регенерационных явлений касается не только внешних проявлений. Существует целый ряд вопросов, носящих проблемный и теоретический характер. К ним относятся вопросы регуляции и условий, в которых протекают восстановительные процессы, вопросы происхождения клеток, участвующих в регенерации, способности к регенерации у различных групп, животных и особенностей восстановительных процессов у млекопитающих.
Установлено, что в конечности амфибий после ампутации и в процессе регенерации происходят реальные изменения электрической активности. При проведении электрического тока через ампутированную конечность у взрослых шпорцевых лягушек наблюдается усиление регенерации передних конечностей. В регенератах увеличивается количество нервной ткани, из чего делается вывод, что электрический ток стимулирует врастание нервов в края конечностей, в норме не регенерирующих.
Попытки стимулировать подобным образом регенерацию конечностей у млекопитающих оказались безуспешными. Так, под действием электрического тока или при сочетании действия электрического тока с фактором роста нервов удавалось получить у крысы только разрастание скелетной ткани в виде хрящевых и костных мозолей, которые не походили на нормальные элементы скелета конечностей.
Несомненна регуляция регенерационных процессов со стороны нервной системы. При тщательной денервации конечности во время ампутации эпиморфная регенерация полностью подавляется и бластема никогда не образуется. Были проведены интересные опыты. Если нерв конечности тритона отвести под кожу основания конечности, то образуется дополнительная конечность. Если его отвести к основанию хвоста — стимулируется образование дополнительного хвоста. Отведение нерва на боковую область никаких дополнительных структур не вызывает. Эти эксперименты привели к созданию концепции регенерационных полей. .
Было установлено, что для инициации регенерации решающим является число нервных волокон. Тип нерва роли не играет. Влияние нервов на регенерацию связывается с трофическим действием нервов на ткани конечностей.
Получены данные в пользу гуморальной регуляции регенерационных процессов. Особенно распространенной моделью для изучения этого является регенерирующая печень. После введения нормальным интактным животным сыворотки или плазмы крови от животных, подвергшихся удалению печени, у первых наблюдалась стимуляция митотической активности клеток печени. Напротив, при введении травмированным животным сыворотки от здоровых животных получали снижение количества митозов в поврежденной печени. Эти опыты могут свидетельствовать как о присутствии в крови травмированных животных стимуляторов регенерации, так и о присутствии в крови интактных животных ингибиторов клеточного деления. Объяснение результатов опытов затрудняется необходимостью учитывать иммунологический эффект инъекций.
Важнейшим компонентом гуморальной регуляции компенсаторной и регенерационной гипертрофии является иммунологический ответ. Не только частичное удаление органа, но и многие воздействия вызывают возмущения в иммунном статусе организма, появление аутоантител и стимуляцию процессов клеточной пролиферации.
Большие разногласия существуют по вопросу о клеточных источниках регенерации. Откуда берутся или как возникают недифференцированные клетки бластемы, морфологически сходные с мезенхимными? Существует три предположения.
1. Гипотеза резервных клеток подразумевает, что предшественниками регенерационной бластемы являются так называемые резервные клетки, которые останавливаются на некоем раннем этапе своей дифференцировки и не участвуют в процессе развития до получения стимула к регенерации.
2. Гипотеза временной дедифференцировки, или модуляции, клеток предполагает, что в ответ на регенерационный стимул дифференцированные клетки могут утрачивать признаки специализации, но затем снова дифференцируются в тот же клеточный тип, т.е., потеряв на время специализацию, они не утрачивают детерминацию.
3. Гипотеза полной дедифференцировки специализированных клеток до состояния, сходного с мезенхимными клетками и с возможной последующей трансдифференцировкой или метаплазией, т.е. превращением в клетки другого типа, полагает, что в этом случае клетка утрачивает не только специализацию, но и детерминацию.
Современные методы исследования не позволяют с абсолютной достоверностью доказать все три предположения. Тем не менее абсолютно верно, что в культях пальцев аксолотля происходит высвобождение хондроцитов из окружающего матрикса и миграция их в регенерационную бластему. Дальнейшая их судьба не определена. Большинство исследователей признают дедифференцировку и метаплазию при регенерации хрусталика у амфибий. Теоретическое значение этой проблемы заключается в допущении возможности или невозможности изменений клеткой ее программы до такой степени, что она возвращается в состояние, когда снова способна делиться и репрограммироватьсвой синтетический аппарат. Например, хондроцит становится миоцитом или наоборот.
Способность к регенерации не имеет однозначной зависимости от уровня организации, хотя давно уже было замечено, что более низко организованные животные обладают лучшей способностью к регенерации наружных органов. Это подтверждается удивительными примерами регенерации гидры, планарий, кольчатых червей, членистоногих, иглокожих, низших хордовых, например асцидий. Из позвоночных наилучшей регенерационной способностью обладают хвостатые земноводные. Известно, что разные виды одного и того же класса могут сильно отличаться по способности к регенерации. Кроме того, при изучении способности к регенерации внутренних органов оказалось, что она значительно выше у теплокровных животных, например у млекопитающих, по сравнению с земноводными.
Регенерация у млекопитающих отличается своеобразием. Для регенерации некоторых наружных органов нужны особые условия. Язык, ухо, например, не регенерируют при краевом повреждении. Если же нанести сквозной дефект через всю толщу органа, восстановление идет хорошо. В некоторых случаях наблюдали регенерацию сосков даже при ампутации их по основанию. Регенерация внутренних органов может идти очень активно. Из небольшого фрагмента яичника восстанавливается целый орган. Об особенностях регенерации печени уже было сказано выше. Различные ткани млекопитающих тоже хорошо регенерируют. Есть предположение, что невозможность регенерации конечностей и других наружных органов у млекопитающих носит приспособительный характер и обусловлена отбором, поскольку при активном образе жизни нежные морфогенетические процессы затрудняли бы существование. Достижения биологии в области регенерации успешно применяются в медицине. Однако в проблеме регенерации очень много нерешенных вопросов.
75.Биологическое и медицинское значение проблемы регенерации. Проявление регенерационной способности у человека. Регенерация патологически измененных органов и обратимость патологически изменённых органов. Регенерационная терапия.
При разрезе в рану устремляется кровь, лейкоциты которой запускают воспалительный процесс. Клетки прилежащей эпителиальной ткани делятся и образуют «струп» (рубец). Потом начинается процесс заживления.
В настоящее время интенсивно изучаются проблемы регенерации, особенно связанные с медициной. Стволовые клетки обладают свойствами:
- стволовая клетка не является окончательно дифференцированной (она скорее детерминирована);
- стволовая клетка способна к неограниченному делению;
- при делении часть клеток остается стволовыми, другая часть подвергается процессу дифференцировки.
Центров по применению стволовых клеток очень мало, в России существует только 2 таких центра. Однако стволовые клетки есть везде. Для лечения и экспериментов берется пуповинная кровь с целью получения стволовых клеток.
Кости черепа в норме не регенерируют. Под руководством И.И.Полежаева происходило удаление участка 10х10 см черепа собаки. Из кости получали путем измельчения костные опилки, которые помещали на рану. В другом эксперименте использовали костные опилки донора и кровь реципиента. Через неделю происходило рассасывание опилок, а к концу 1 года рана зарастала.
Большое значение имеет регенерация после радиоактивного облучения. Малые дозы стимулируют, а большие, наоборот ингибируют данный процесс.
Если провести механическое раздавливание культи или помещение ее в кислоту – регенерация идет в 50% случаев.
Елизаров проводил ломку и удлинение костей. Им были созданы уникальные аппараты, благодаря которым было возможно раздвижение костей скелета и коррекция их формы.
Остро стоит проблема регенерации печени. При циррозе печени приходится проводить ее частичное удаление. Иногда подобная операция проводится несколько раз, печень быстро регенерирует без сохранения формы, сохраняя функцию и общую массу.
Регенерацию можно стимулировать антикейлоном, витамином В12, АТФ, РНК.
Выделяют типы регенерации в патологически измененных органах.
- Регенерация после воздействия токсических веществ.
- Регенерация после воздействия вредных физических факторов.
- Регенерация после заболеваний, вызываемых микроорганизмами и вирусами.
- Регенерация после нарушения кровоснабжения.
- Регенерация после голода, гипокинезии (обездвиживании), атрофии.
- Регенерация после повреждений, вызываемых в организме нарушением функции органов.
76.Понятие о гомеостазе. Общие закономерности гомеостаза живых систем. Генетические, клеточные и системные основы гомеостатических реакций организма. Роль эндокринной, нервной и иммунной систем в обеспечении гомеостаза и адаптивных изменений.
Термин «гомеостаз» был предложен для понимания постоянства состава лимфы, крови и тканевой жидкости. Гомеостаз характерен для любой системы, это своего рода обобщение множества частных проявлений стабильности системы.
Как же организм сохраняет постоянство??
Гомеостаз – поддержание постоянства внутренней среды организма в непрерывно изменяющихся условиях внешней среды. Т.к. организм – многоуровневый саморегулирующийся объект, его можно рассматривать с точки зрения кибернетики. Тогда, организм – сложная многоуровневая саморегулирующаяся система с множеством переменных.
Переменные входа:
- причина;
- стимул;
- раздражение.
Переменные выхода:
- эффект;
- ответ;
- реакция;
- следствие.
Причина – отклонение от нормы реакции в организме. Решающая роль принадлежит обратной связи. Существует положительная и отрицательная обратная связь.
Отрицательная обратная связь уменьшает действие входного сигнала на выходной. Положительная обратная связь увеличивает действие входного сигнала на выходной эффект действия.
Живой организм – ультрастабильная система, осуществляющая поиск наиболее оптимального устойчивого состояния, которое обеспечивается адаптациями.
Адаптация – поддержание переменных показателей на поведенческом, анатомическом, биохимическом и других уровнях.
Этология – наука, изучающая поведение животных и человека. Типы поведения животных и человека ограничены их морфологическими и физиологическими особенностями. У человека есть зависимость поведения от типа сложения. Существуют 3 типа сложения:
эндоморфный;
эктоморфный;
мезоморфный.
Животные могут совершенствовать свои движения за счет информации, кроме того, они имеют возможность регулировать их. Животные должны различать объекты внешней среды, получать информацию при помощи органов чувств. Полученная информация подвергается переработке нервной и эндокринной системами. Многие типы поведения могут вызывать гормональные изменения.
Морфологические и физиологические признаки подвержены естественному отбору, поведение в свою очередь, зависит от этих признаков, а значит, зависит и от естественного отбора. Поведение передается по наследству, повышает приспособляемость, увеличивает продолжительность жизни, количество потомков. Различные поведенческие реакции позволяют использовать благоприятные условия среды, защищают организм от неблагоприятных условий. Например, у пчел поддержание чистоты в улье. За гигиеническое поведение отвечают как минимум 2 гена. Поддержание чистоты защищает пчел от болезней. Поведение ящерицы, отбрасывающей хвост, если это необходимо, - тоже приспособительная реакция. Другие типы поведенческих реакций наблюдаются при защите от хищников, при поиске пищи, партнера, защите потомства и многих других случаях. Некоторые насекомые выделяют особые химические вещества – феромоны для привлечения потомства. В брачный период лягушки квакают и их «песня» видоспецифична.
Поведенческие признаки обладают не только адаптивными свойствами, но могут также и наследоваться, что обусловливает естественный отбор. Не все типы поведения получаются при передаче с генами, они могут приобретаться – благоприобретенные. Резкую границу между теми и другими провести нельзя, т.к. гены и среда тесно взаимодействуют друг с другом, поэтому выделить отдельно генетические и благоприобретенные свойства нельзя.
Можно привести следующие примеры генетических свойств. Хорея Гентингтона – наследственное заболевание, «танец», поражает ЦНС, у больных также нарушена пространственная ориентация. Другой пример, дауны бывают благожелательными, ласковыми, подражают действиям здоровых людей
Итак, важные свойства поведенческих реакций:
- поведение подвержено действию естественного отбора;
- поведенческие признак возникают из анатомии, морфологии и физиологии животного неотделимы о них;
- формы поведения обычно адаптивны и часто могут передаваться либо генетически, либо в результате обучения;
- у многих биологических видов существуют определенные формы поведения.
Если организм не смог адаптироваться на поведенческом уровне, он делает это на биохимическом уровне. Биохимическая адаптация очень сложна, наиболее характерна для растений, т.к. животному проще мигрировать.
Процесс адаптации бывает по времени:
- эволюционная адаптация;
- акклиматизация;
- немедленная адаптация.
Эволюционная адаптация – длительный процесс, приобретение новой генетической информации, изменяется генотип, следовательно, изменяется и фенотип. Для своего завершения подобная адаптация требует многих поколений.
Акклиматизация – адаптации, которые происходят в процессе жизни в естественных условиях.
Акклимация– адаптации, происходящие в искусственных условиях.
Происходит в течение нескольких часов – лет (зима – лето). Смена часовых поясов, перевод времени.
Немедленная адаптация сопровождается почти мгновенной адаптивной реакцией (психогенное воздействие, переход из тепла в холод). Кратковременная реакция.
Любая адаптация возникает в результате взаимодействия генетических факторов и факторов внешней среды.
Генетический аспект гомеостаза рассматривают с 3 позиций:
- гомеостаз генотипа;
- гомеостаз организма как целого. Контроль за единством генотипа всего организма. Поддержание гомеостаза осуществляется при гибели видоизмененных клеток.
- гомеостаз популяции. Закон генетической стабильности в популяции.
В поддержании гомеостаза участвуют различные системы.
Нервная сигнализация – основной инструмент передачи и оценки сигналов из внутренней и внешней среды.
Гормоны принимают участие в регуляции гомеостаза. Регулируют обмен веществ, воды, белков, липидов, углеводов, энергии, электролитов. Контролируют работу всех органов, в том числе почек, печени, ЦНС.
Иммунная система защищает постоянство внутренней среды организма от факторов 2-х групп:
- микроорганизмов и экзогенных факторов с признаками чужеродной генетической информацией;
- соматических мутаций. Достаточно изменений в 1-2 генах, чтобы сработала иммунная система.
77.Проблемы трансплантации органов и тканей. Ауто-, алло- и ксенотрансплантация, трансплантация жизненно важных органов. Иммунитет. Тканевая несовместимость и пути ее преодоления. Искусственные органы.
Из-за бурного развития трансплантологии остро встал вопрос о трансплантационном иммунитете.
Трансплантология – медико-биологическая наука, изучающая вопросы заготовки, консервирования и пересадки органов и тканей.
Трансплантационный иммунитет – своеобразная реакция организма на трансплантацию, проявляющаяся в отторжении пересаженных органов и тканей.
Дата добавления: 2015-04-18; просмотров: 308; Мы поможем в написании вашей работы!; Нарушение авторских прав |