КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ГЛАВА 3.

|
| Рис. 3.21. Выключение транскрипции бактериального оперона. Связывание корепрессора (С) с неактивным репрессором 0R) изменяет конформацию последнего. Комплекс корепрессор— репрессор (С— IR) связывается с оператором и блокирует транскрипцию. |
ляется их транскрипция, поскольку репрессорный белок неактивен. В этом случае специфический эффектор (корепрессор), связываясь с неактивным репрессором, вызывает в нем такие конформационные изменения, которые обеспечивают связывание комплекса с операторным участком, и транскрипция оперона выключается (рис. 3.21). Сам по себе репрессор не способен связываться с оператором, поэтому при уменьшении концентрации корепрессора транскрипция возобновляется.
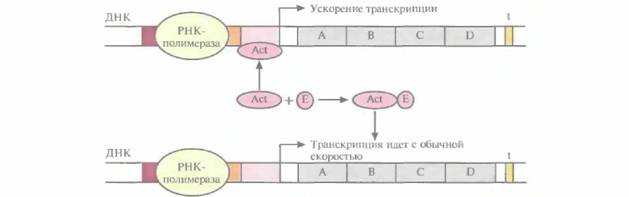
Регуляция транскрипции с помощью репрессора называется отрицательной. Если же система регуляции направлена на повышение скорости транскрипции, то она называется положительной Рассмотрим вкратце этот процесс. Белок-активатор связывается с участком между ТАТА-боксом и сайтом инициации транскрипции. При этом он не
|
| Рис. 3.22. Изменение скорости транскрипции. Активатор (Act) связан с участком между ТАТА-боксом и сайтом инициации транскрипции, скорость транскрипции повышена. Эффектор (Е) связывается с активатором и препятствует его соединению с ДНК; скорость транскрипции уменьшается. |
ДНК,РНК и синтез белка 45

|
| Структура дезоксирибонуклеиновой кислоты J.D. Watson, F.H.C. Crick Nature 171; 737-738, 1953 |
| Выяснение молекулярной структуры генетического материала -ДНК — без сомнения стало одним из самых замечательных научных достижений XX в. Уотсон и Крик описали свое открытие так: «Мы хотим предложить структуру соли дезоксирибонуклеиновой кислоты (ДНК). Эта структура обладает весьма необычными свойствами. представляющими большой био-логический интерес... Она образована двумя спиральными цепочками, закрученными вокруг общей оси... Обе спирали правые, но... последовательности атомов в них взаимно противоположны... Весьма интересен способ, с помощью которого цепочки удерживаются вместе... Пуриновые и пиримидиновые основания образуют пары, при этом пуриновое основание одной цели соединяется водородными связями с пиримидиновым основанием другой... Если одно из оснований пары — это аденин, то... вторым основанием должен | быть тимин; то же самое относится к гуанину и цитозину. Последовательность оснований в полинуклеотидной цепи может быть любой... Очень важно, что из предложенного нами механизма специфического спаривания непосредственно вытекает возможность копирования генетического материала». В следующей статье, опубликованной несколькими месяцами позже (Nature 171: 964, 1953), Уотсон и Крик уточняют: «Сахарофосфатный остов в нашей модели абсолютно постоянен, но в эту структуру может вписаться любая последовательность пар нуклеотидов... В длинной молекуле возможно безграничное число перестановок, и нам кажется вполне вероятным, что точная последовательность оснований содержит в закодированном виде генетическую информацию... Наша модель дает объяснение многим феноменам. Например, спонтанная | мутация может возникнуть в результате случайного перехода одного из оснований в редкую таутомерную форму, а образование пар гомологичных хромосом при мейозе может обусловливаться специфическим спариванием оснований». За десять лет, прошедших после обнародования теории двойной спирали ДНК и принципа комплементарности, раскрыты молекулярные механизмы репликации ДНК; установлены процессы, отвечаюшие за расшифровку генетической информации и регуляцию синтеза генных продуктов; выяснены многие причины, по которым эти продукты синтезируются в измененном виде. Со времени выхода в свет этой публикации и до наших дней открытие Уотсона и Крика нисколько не утратило своего значения, В частности, если бы не была установлена структура ДНК, сейчас не существовало бы технологии рекомбинантньгх ДНК. |
только не блокирует перемещение РНК-полимеразы вдоль молекулы ДНК, а напротив, ускоряет его, действуя как своего рода «смазка». Активаторы специфичны для определенных сайтов активации. Иногда с работающим активатором связывается эффектор, переводящий его в неактивную форму; тогда скорость транскрипции уменьшается (рис. 3.22). В других случаях эффектор, напротив, активирует неработающий активатор. Чтобы понять детали регуляции транскрипции у бактерий, необходимо провести тщательный анализ мутаций, которые влияют на данную регуляторную систему, и исследовать in vitro различные сайты связывания белков и ДНК.
Регуляция транскрипции у эукариот
Некий набор основных структурных генов, обеспечивающих жизнедеятельность клетки, — генов «домашнего хозяйства» — транскрибируется в большинстве активно функционирующих эукариотических клеток. В отличие от этого специфические гены, которые отвечают за уникальность тех или иных тканей или органов, транскрибируются и транслируются только в определенных клетках. Так, гены, кодирующие a- и ß-субъединицы гемоглобина взрослого человека, экспрессируются исключительно в клетках — предшественниках эритроцитов. Число разных мРНК, специфичных для разных клеток, варьирует от единиц до десятков. Способность клеток включать (активировать) или выключать (ингибировать) структурные гены крайне важна для поддержания клеточной специфичности и экономного расходования энергетических ресурсов.
Для включения и выключения транскрипции различных эукариотических структурных генов используется множество разнообразных высоко-
46 ГЛАВА 3

|
| Рнс. 3.23. Регуляторные элементы структурных генов эукариот. Знаки «— » при числах означают, что эти элементы находятся в молекуле ДНК слева от сайта инициации транскрипции (+1). Стрелка — направление транскрипции. Положение регуляторных элементов и их размер даны без соблюдения масштаба. |
специфичных процессов. Но так или иначе регуляция транскрипции у эукариот осуществляется с помощью специфических белков -- факторов транскрипции. Многие из них связываются непосредственно с нуклеотидной последовательностью длиной менее 10 п.н., называемой по-разному: боксом, модулем, элементом инициации, регуляторным элементом. В отличие от прокариот у эукариот опероны в большинстве своем отсутствуют, т. е. каждый эукариотический структурньй ген имеет свой собственный набор регуляторных элементов. Существенную роль в регуляции транскрипции у эукариот, помимо опосредованной взаимодействием между ДНК и белками, играют также белок-белковые взаимодействия.
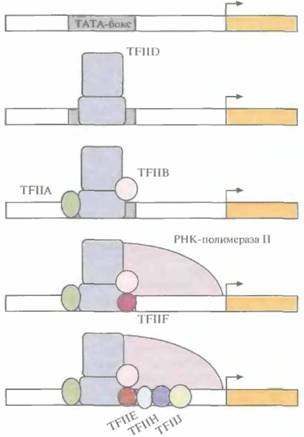
Несмотря на индивидуальность набора регуляторных элементов у структурных генов эукариот, каждый из них имеет промоторный участок (ТАТА-бокс, или бокс Хогнесса) из восьми нуклеотидов, включающий последовательность ТАТА; последовательность ССААТ (САТ-бокс); участок из повторяющихся динуклеотидов GC (GC-бокс). Эти элементы находятся на расстоянии 25, 75 и 90 п.н. от сайта инициации соответственно (рис. 3.23). Транскрипция структурного гена эукариот начинается со связывания с ТАТА-боксом фактора транскрипции HD (TFIID), который представляет собой комплекс по крайней мере из 14 белков. Затем с TFIID и участками ДНК, примыкающими к ТАТА-бок-су, связываются другие факторы транскрипции, и, наконец, со всем этим транскрипционным комплексом связывается РНК-полимераза II.Затем при участии дополнительных факторов происходит инициация транскрипции в точке + 1 (рис. 3.24). Ясно, что если последовательность ТАТА отсутствует или существенно изменена,
|
| Рис, 3.24. Инициация транскрипции структурного гена эукариот. Сначала фактор транскрипции TFIID связывается с ТАТА-боксом, затем происходит присоединение других факторов транскрипции и РНК-полимеразы II и, наконец, вспомогательных факторов, запускающих транскрипцию. Стрелка направление транскрипции. |
то транскрипция структурного гена становится невозможной. Идентифицированы также факторы транскрипции, специфичные для регуляторных элементов ССААТ и GC, но пока неясно, как ДНК-белковые взаимодействия могут влиять в этом случае на эффективность транскрипции, если элементы расположены на расстоянии более 75 п.н. от сайта инициации. Кроме того, на расстоянии сотен и даже тысяч пар оснований от сайта инициации находится так называемая энхансерная последовательность, которая многократно повышает скорость транскрипции структурных генов. По-видимому,
ДНК, РНК и синтезбелка 47
сближение удаленных регуляторных элементов и соответствующего структурного гена происходит при укладке хромосомной ДНК. Кроме того, факторы транскрипции, которые связываются с определенными энхансерами и регуляторными элементами, могут образовывать цепочку, соединяющую удаленные друг от друга сайты.
Некоторые репрессированные (неэкспрессирующиеся) гены активируются каскадом событий, который запускается каким-либо специфическим внеклеточным сигналом, например повышением температуры или синтезом гормона. Гормон, поступив в кровоток, связывается с рецепторами специфических клеток, облегчающими его проникновение в клетку. Оказавшись в клетке, гормон вступает во взаимодействие с одним из клеточных белков и изменяет его конформацию. В таком измененном состоянии белок проникает в ядро и связывается со специфическим регуляторным элементом, который инициирует транскрипцию соответствующего гена.
Существуют также белки, которые, взаимодействуя с регуляторными элементами, блокируют транскрипцию. Например, известен класс генов позвоночных (примерно 18), активно транскрибирующихся только в нервных клетках. Каждый из этих генов имеет регуляторный элемент из 24 п.н., находящийся «левее» (upstream) сайта +1; он обозначается NRSE (от англ, neuron-restrictive silencer element). Во всех клетках, кроме нейронов, синтезируется NRSF-фактор (от англ, neuron-restrictive silencer factor), который связывается с NRSE и блокирует транскрипцию соответствующих генов. В нейронах NRSF не синтезируется, и упомянутые гены активно транскрибируются.
Итак, регуляция транскрипции у эукариот -это очень сложный процесс. Структурный ген может иметь множество регуляторных элементов, которые активируются специфическими сигналами в клетках разного типа в разное время клеточного цикла. Однако некоторые структурные гены находятся под контролем уникального фактора транскрипции. Специфические белки могут взаимодействовать с определенными регуляторными элементами и блокировать транскрипцию или связываться со всем транскрипционным комплексом еще до инициации транскрипции или во время элонгации.
ЗАКЛЮЧЕНИЕ
Молекула ДНК состоит из двух антипараллельных полинуклеотидных цепей, образующих двойную спираль. Их мономерной единицей является нуклеотид, который состоит из азотистого основания, дезоксирибозы и фосфатной группы. Соседние нуклеотиды в цепи связаны фосфодиэфирными связями, а цепи удерживаются вместе с помощью водородных связей, образующихся между комплементарными основаниями. При этом аденин образует водородные связи только с тимином, гуанин — только с цитозином. Процесс удвоения ДНК называется репликацией. В нем участвует множество различных белков, прежде всего ДНК-полимеразы. Каждая из цепей ДНК служит матрицей для синтеза комплементарной цепи. Комплемен-тарность оснований противоположных цепей гарантирует идентичность новосинтезированной и исходной ДНК.
Ключевую роль в осуществлении всех биологических функций играют белки. Белковая молекула — это полипептид, состоящий из аминокислот, которые соединены друг с другом пептидными связями. Последовательность аминокислот в белке определяется последовательностью нуклеотидов в ДНК. В синтезе белков участвуют молекулы РНК (мРНК, рРНК и тРНК), различные ферменты и белковые факторы. Все РНК синтезируются на ДНК как на матрице; этот процесс называется транскрипцией. Правильность транскрипции, т. е. ее начало и завершение в нужных сайтах, обеспечивают специфические нуклеотидные последовательности в ДНК и белковые факторы. У эукариот большинство структурных генов состоит из кодирующих (экзоны) и некодирующих (интроны) участков. Первичные транскрипты содержат как те, так и другие. Однако по окончании транскрипции интроны вырезаются, а экзоны сшиваются с образованием функциональной мРНК. В мРНК содержится в закодированном виде информация о последовательности аминокислот в молекуле соответствующего белка.
Синтез белка называется трансляцией. Важную роль в нем играют молекулы тРНК и рРНК. В клетке присутствует более 50 разных тРНК.
Дата добавления: 2015-04-16; просмотров: 282; Мы поможем в написании вашей работы!; Нарушение авторских прав |