КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ГЛАВА 8 3 страница
Применение моноклональных антител позволяет существенно повысить специфичность метода ELISA, поскольку они связываются с одним, строго определенным антигенным сайтом. К настоящему времени получен целый ряд моноклональных антител, которые можно использовать для обнаружения различных соединений и патогенных микроорганизмов (рис. 9.3). Альтернативой получению моноклональных антител из культуры гибридомных клеток может быть отбор и производство моноклональных антител и их частей (Fv-фрагментов), направленных против антигена-мишени, с помощью Е. coli (см. гл. 10).
Системы ДНК-диагностики
Информация о всем многообразии свойств организма заключена в его генетическом материале. Так, патогенность бактерий определяется наличием у них специфического гена или набора генов, а наследственное генетическое заболевание возникает в результате повреждения определенного гена. Сегмент ДНК, детерминирующий данный биологический признак, имеет строго определенную нуклеотидную последовательность и может служить диагностическим маркером.
В основе многих быстрых и надежных диагностических методов лежит гибридизация нуклеиновых кислот — спаривание двух комплементарных сегментов разных молекул ДНК. Процедура в общих чертах состоит в следующем.
1. Фиксация одноцепочечной ДНК-мишени на мембранном фильтре.
2. Нанесение меченой одноцепочечной ДНК-зонда, которая при определенных условиях (температуре и ионной силе) спаривается с ДНК-мишенью.
3. Промывание фильтра для удаления избытка несвязавшейся меченой ДНК-зонда.
4. Детекция гибридных молекул зонд/мишень.
В диагностических тестах, основанных на гибридизации нуклеиновых кислот, ключевыми являются три компонента: ДНК-зонд, ДНК-мишень и метод детекции гибридизационного сигнала. Система детекции должна быть в высшей степени специфичной и высокочувствительной.

|
| Рис. 9.3. Использование моноклональных антител для выявления различных соединений и диагностики инфекционных заболеваний. |
188 ГЛАВА 9
Гибридизационные зонды
Чтобы обеспечить адекватность диагностического теста, гибридизационные ДНК- и РНК-зонды должны быть высокоспецифичными. Другими словами, необходимо, чтобы зонд гибридизовался только с искомой нуклеотидной последовательностью. Если есть вероятность получения ложноположительного (наличие гибридизационного сигнала в отсутствие последовательности-мишени) или ложноотрицательного (отсутствие сигнала при наличии последовательности-мишени) результата, то целесообразность применения теста значительно снижается. Специфичность зондов может проявляться на разных уровнях: они могут «различать» два и более вида, отдельные штаммы в пределах одного вида или разные гены. В зависимости от ситуации зонды могут быть представлены молекулами ДНК или РНК; они могут быть длинными (более 100 нуклеотидов) или короткими (менее 50 нуклеотидов), представлять собой продукт химического синтеза, клонированные интактные гены или их фрагменты.
Зонды получают разными способами. Один из них состоит в следующем, ДНК патогенного микроорганизма расшепляют с помощью рест-ринируюшей эндонуклеазы и клонируют в плазмидном векторе. Затем проводят скрининг рекомбинантных плазмид с использованием геномной ДНК как патогенного, так и непатогенного штаммов. Те плазмиды, которые содержат последовательности, гибридизующиеся только с ДНК патогенного штамма, составляют основу вилоспепифичных зондов. После этого проводят ряд дополнительных гибридизаций с ДНК, выделенными из различных организмов, чтобы удостовериться, что потенциальные зонды не дают с ними перекрестной гибридизации. Для определения чувствительности метода каждый из зондов проверяют также на модельных образцах, в том числе и на смешанных культурах.
Весьма желательно, чтобы ДНК-диагностику можно было проводить на исходном материале, без дополнительного его культивирования или выделения нуклеиновых кислот, особенно в тех случаях, когда тестируются клинические образцы. Исследователи с успехом проводят гибридизацию с ДНК-мишенями, присутствующими в образцах кала, мочи, крови, смывах из зева и в тканях без предварительной их очистки. Если концентрация последовательности-мишени в исследуемом образце слишком мала, ее можно амплифицировать с помощью полимеразной цепной реакции (ПЦР).
Диагностика малярии
В качестве примера использования ДНК-зондов для диагностики заболеваний можно привести процедуру обнаружения Plasmodium falciparum. Этот паразит вызывает малярию, заболевание, угрожающее примерно трети всего населения Земли. Он инфицирует эритроциты и разрушает их, что приводит к развитию лихорадки, а в тяжелых случаях — к поражению мозга, почек и других органов. Чтобы выявить источники инфекции, оценить эффективность мер по их ликвидации и обеспечить раннюю диагностику и лечение, необходимы достаточно чувствительные, простые и недорогие методы. В настоящее время малярию диагностируют с помощью микроскопического исследования мазков крови -эффективного, но трудоемкого и занимающего много времени процесса. Иммунологические методы обнаружения Plasmodium, такие как ELISA, достаточно быстрые и их легко автоматизировать, но с их помощью нельзя отличить текущую инфекцию от прошедшей, поскольку при этом определяется только наличие антител к Plasmodium в крови больных.
Для избирательной ДНК-диагностики текущей инфекции, т. е. для выявления ДНК возбудителя, в качестве основы используются высокоповторяющиеся последовательности ДНК P. falciparum. Сначала с помощью ДНК-зонда проводится скрининг библиотеки геномной ДНК паразита. Затем отбираются клоны, дающие наиболее интенсивный гибридизационный сигнал, поскольку именно они предположительно содержат высокоповторяющиеся последовательности. ДНК каждого из отобранных клонов проверяют на способность к гибридизации с ДНК видов Plasmodium, не вызывающих малярию. В качестве специфического зонда выбирается последовательность, гибридизующаяся с ДНК Р. falciparum, но не с ДНК P. vivax, P. cynomolgi или с ДНК человека. С его помощью
Молекулярная диагностика 189

| |
| Выявление аллелей ß-глобиноиого гена методом гибридизации с синтетическими олигонуклеотидами | |
| В. J. Conner. A. A. Reyes, С. Morin, K. Itakura, R. L. Teplitz. and R, B. Wallace Proc, Natl. Atad. Sei. VSA 80: 278-282. )983 |
| С разработкой в начале 80-х гг. быстрых, эффективных и недорогих методов химического синтеза олигонуклеотидов появилась возможность использовать радиоактивно меченные олигонуклеотидные зонды для выявления различий между нуклеотидными последовательностями, в частности для обнаружения мутации у человека. К тому времени было изолировано относительно небольшое число генов, главным образом в виде комплементарных ДНК (кДНК), поэтому подбор зонда, гомологичного конкретному гену, представлял собой непростую задачу. Но даже если соответствующие кДНК были получены, невозможно было различить нормальный и мутантный гены чело- | века, отличающиеся друг от друга лишь одной парой оснований, проводя гибридизацию с протяженным (более 100 п. н.) зондом. В 1983 г. Коннер и др. синтезировали специфичные олигонуклеотиды. способные распознавать нормальную и мутантную ДНК, при этом можно было установить, гомозиготен или гетерозиготен данный индивидуум по исследуемому гену. Для диагностики серповидноклеточной анемии они использовали два олиго-нуклеотида длиной по 19 п. н.; один был комплементарен нормальному аллелю β-глобинового гена (βA), а другой — мутантному (ßs). ДНК здорового человека (βAρΑ) гибридизовалась только с βA-зондом, ДНК больного серпо- | видноклеточной анемией (ßsßs) — только с ßs-зондом, а ДНК индивидуума, гетерозиготного по данному гену (ßAßs), — с обоими зондами. Эта модельная система впервые продемонстрировала возможность определения генотипов с помощью гибридизации н положила начало молекулярной диагностике многих генетических заболевании человека. Вслед за этим был разработан целый ряд ДНК-тестов для выявления мутаций. И хотя гибрид и за ц ионный метод сейчас уступил место более совершенным методам, таким как ПЦР и ЛОЗ (лигирование олигонуклеотидных зондов), он сыграл свою роль, проиллюстрировав возможность определения точковых мутаций. | |||
можно обнаруживать всего 10 пг очищенной ДНК P.falcipantm или 1 нг той же ДНК в крови больного.
Получены и охарактеризованы более 100 различных ДНК-зондов, позволяющих обнаруживать патогенные штаммы различных бактерий, вирусов и паразитических простейших. Так, имеются зонды для диагностики бактериальных, инфекций человека, вызываемых Legionella рпеи-mophïla (респираторные заболевания). Salmonella typhi {пищевые отравления), Campylobacter hyoin-testinaiis (гастриты), а также для выявления энте-ротоксичного штамма Escherich'ia colî (гастроэнтериты). Однако это лишь «верхушка айсберга»; в принципе с помощью гибридизации можно выявлять практически любые патогенные микроорганизмы.
Выявление Trypanosoma cruzi
Паразитическое простейшее Tiypanosorna cruzi вызывает болезнь Чагаса (южноамериканский трипаносомоз), уносящую ежегодно примерно 50 тыс. жизней. Паразит широко распространен в Латинской Америке. Он переносится клопами-хищнецами, проникает в печень, селезенку, лимфатические узлы, центральную нервную систему и, размножаясь, разрушает клетки, в которых паразитирует. Для диагностики острой формы болезни Чагаса обычно проводят микроскопическое исследование свежей пробы периферической крови. Можно использовать и другой тест, более длительный, но выявляющий паразитов с большей вероятностью. Незараженных насекомых кормят кровью пациента и через 30—40 суг исследуют под микроскопом их кишечник на предмет наличия паразитов. Оба метода весьма трудоемки, дорогостоящи и требуют длительного времени. Болезнь можно диагностировать и иммунологическими методами, однако они часто дают ложноположительные результаты. В качестве альтернативы этим менее чем удовлетворительным процедурам было разработано несколько подходов, основанных на применении ПЦР. В насто-
190 ГЛАВА 9
ящее время ПЦР-диагностика болезни Чагаса служит дополнением к традиционным, широко используемым методам.
Один из ПЦР-тестов основан на выявлении фрагмента ДНК длиной 188 п. н., который присутствует во множестве копий в геноме Т. cruzi, но отсутствует в геномной ДНК нескольких родственных паразитов. После амплификации этот фрагмент без труда обнаруживается с помощью электрофореза в полиакриламид-ном геле. Незначительно варьируя методику проведения ПЦР (например, изменяя нуклеотидную последовательность праймеров), последнюю можно использовать для обнаружения широкого спектра бактерий, вирусов и паразитов.
Нерадиоактивные методы детекции
В большинстве лабораторий для гибридизации используют зонды, меченные каким-либо радиоактивным изотопом, чаще всего 32Р. Такие зонды обладают высокой удельной радиоактивностью и обеспечивают хорошее отношение сигнал/шум. Радиоактивно меченный зонд наносят на фильтр с фиксированной на нем ДНК-мишенью, проводят гибридизацию, отмывают несвязавшуюся ДНК-зонд и детектируют метку с помощью радиоавтографии.
Однако 32P является короткоживущим изотопом, испускающим высокоэнергетическое излучение; при работе с ним необходимо использовать специальное оборудование и обеспечивать безопасную утилизацию отходов. Чтобы обойти эти трудности, были созданы нерациоактивные системы детекции. Для усиления гибридизаци-онного сигнала в этом случае используется ферментативное превращение хромогенного или хемилюминесцентного субстрата: первый из них под действием фермента изменяет окраску, а второй испускает свет. В большинстве подобных систем применяются ДНК-зонды, содержащие биотинилированные нуклеотиды. Гибридизация и детекция сигнала проводятся более или менее стандартным образом.
1. Зонд, меченный биотином, гибридизуют с ДНК-мишенью (рис. 9.4, А).
2. Промывают фильтр для удаления избытка несвязавшегося зонда.
3. Добавляют авидин (белок куриного яйца) или стрептавидин (бактериальный аналог авидина) (рис. 9.4, Б).
4. Добавляют биотинилированный фермент щелочную фосфатазу или пероксидазу хрена (рис. 9.4, В).
5. В зависимости от используемого фермента добавляют хромогенный или хемилюминесцентный субстрат и регистрируют изменение окраски либо люминесценцию, сопровождающую превращение субстрата в продукт (рис. 9,4, Г).
В качестве альтернативы после гибридизации ДНК с биотинилированным зондом можно добавлять уже готовый комплекс стрептавидин—фермент, имеющий сайт связывания с биотином.
Как авидин, так и стрептавидин связываются с биотином очень прочно (константа диссоциации (Кd = 10~15); кроме того, каждый из белков имеет четыре независимых биотинсвязывающих сайта, благодаря чему одна молекула авидина или стрептавидина может одновременно присоединять фермент и зонд, меченные биотином. Биотинилирование и связывание со стрептавидином не приводят к снижению ферментативной активности. В хромогенных системах детекции в том месте, где находится гибридная ДНК, под действием фермента образуется нерастворимый краситель, а в хемилюминесцентных системах — продукт, который испускает свет.
Нерадиоактивные системы детекции обладают и другими преимуществами: биотинилированная ДНК остается стабильной при комнатной температуре как минимум год; методы регистрации хемилюминешенции обладают такой же чувствительностью, как и методы регистрации радиоактивного сигнала; детекция испускаемого света при помощи рентгеновской пленки или люминометра, как и регистрация изменения цвета, занимают несколько часов. По-видимому, хемилюминесцентные системы регистрации сигнала, все же более чувствительные, чем хромогенные, вскоре вытеснят все остальные, использующиеся при ДНК-диагностике. Если при этом применяется ГЩР, то амплифицируемый продукт можно пометить флуоресцентным красителем, присоединяя его
Молекулярная диагностика 191
| Рис. 9.4. Хемилюминесцентный метод обнаружения ДНК-мишени. Б — биотип, ЩФ — щелочная фосфатаза. А. Связывание биотинилированного зонда с ДНК-мишенью. В. Связывание стрептавидина с биотином. В. Связывание биотинилированной щелочной фосфатазы со стрептавидином. Г. Образование люминесцирующего продукта под действием щелочной фосфатазы. | 
|
к 5'-концу каждого праймера. В качестве красителей часто используют флуоресцеин и родамин, которые испускают зеленый и красный свет соответственно. После ПЦР-амплификации ДНК-мишени проводят разделение флуоресцеин-ме-ченного праймера и продуктов амплификации, после чего регистрируют включение метки (рис. 9.5), Если ДНК-мишень в образце отсутствует, то не будет образовываться и флуоресцирующий продукт.
Один из недавно разработанных нерадиоактивных методов детекции основан на использовании зонда — «молекулярного маяка» (рис. 9.6). Такой зонд состоит из 25 нуклеотидов. Средние 15 из них комплементарны ДНК-мишени и не спариваются друг с другом, а 5 концевых нуклеотидов взаимно комплементарны и образуют шпильку. К 5'-концу присоединен флуоресцентный хромофор (флуорофор), а к 3'-концу — нефлуоресцентный хромофор (тушитель), на который передается энергия возбуждения флуорофора. В растворе при комнатной температуре «маяк» имеет такую конфигурацию, при которой флуорофор и тушитель находятся в тесном контакте, и флуоресценция флуорофора тушится. Когда же 15 средних нуклеотидов зонда гибридизуются с комплементарной последовательностью ДНК- или РНК-мишени, происходит пространственное разделение
192 ГЛАВА 9

| Рис. 9.5. Детекция ПЦР-продуктов с помощью флуоресцентного красителя, присоединенного к праймерам (Р1 и Р2), |
флуорофора и тушителя, и зонд испускает свет. Температура реакционной смеси должна быть близка к комнатной, поскольку при ее повышении шпилька денатурирует, флуорофор и тушитель расходятся и происходит флуоресценция. Необходимо также, чтобы все 15 нуклеотидов зонда были комплементарны соответствующей последовательности ДНК- или РНК-мишени.
Геномная дактилоскопия
Метод геномной дактилоскопии (ДНК-типирование) часто используется в судебной медицине для идентификации биологических образцов. С его помощью можно доказать, что подозреваемый действительно совершил преступление, или, напротив, что он невиновен. Для проведе-
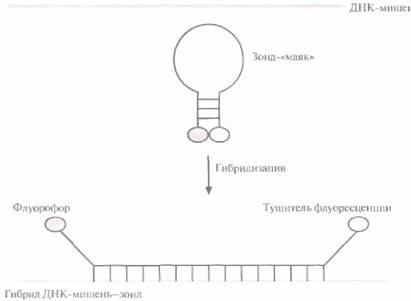
| 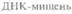 Рис. 9.6. Гибридизация зонда — «молекулярного маяка» с ДНК-мишенью. Одноцепочечная область зонда гибридизуется с комплементарной последовательностью ДНК-мишени, его «шпилька» разрушается, флуорофор и тушитель флуоресценции перестают контактировать друг с другом и наблюдается флуоресценция. Это указывает на то, что между зондом и последовательностью-мишенью произошла гибридизация. Из работы Tyagi, Kramer, Nat. Biotechnol. 14: 303-308, 1996, с изменениями
Рис. 9.6. Гибридизация зонда — «молекулярного маяка» с ДНК-мишенью. Одноцепочечная область зонда гибридизуется с комплементарной последовательностью ДНК-мишени, его «шпилька» разрушается, флуорофор и тушитель флуоресценции перестают контактировать друг с другом и наблюдается флуоресценция. Это указывает на то, что между зондом и последовательностью-мишенью произошла гибридизация. Из работы Tyagi, Kramer, Nat. Biotechnol. 14: 303-308, 1996, с изменениями
|
Молекулярная диагностика 193
ния ДНК-типирования сначала берут часть биологического образца (пробу крови, сперму, кусочек кожи, волосы) и определяют, достаточно ли в нем интактной ДНК для последующего анализа. Затем ДНК подвергают эндонуклеазному расщеплению, полученные фрагменты разделяют в агарозном геле и переносят на най-лоновый фильтр. Проводят последовательную гибридизацию с четырьмя или пятью радиоактивно меченными зондами, каждый из которых распознает определенную последовательность ДНК (при этом перед гибридизацией со следующим зондом предыдущий полностью удаляют с мембраны). После каждой гибридизации с помощью радиоавтографии визуализируют полосы, отвечающие продуктам гибридизации зонда с рестрицированной ДНК, и строят «лестницу фрагментов» для всех образцов (рис. 9.7). Каждый этап (гибридизация и радиоавтография) длится от 10 до 14 сут, так что вся процедура может занять много недель и даже несколько месяцев. В качестве зондов обычно используют минисателлитные ДНК, многократно встречающиеся в геноме человека и состоящие из тандемно повторяющихся участков. Длина повторов варьирует от 9 до 40 п. н., а их число — от 10 до 30; при этом одни и те же минисателлитные последовательности у разных индивидов могут иметь разную длину. Зги различия возникают в результате увеличения или уменьшения числа тандемных повторов, по-видимому, в ходе репликации ДНК. Никаких биологических последствий такие вариации не имеют, поскольку минисателлитные ДНК не кодируют белков. Ребенок наследует одну минисателлитную последовательность от одного родителя, а другую — от другого.
«ДНК-отпечаток" данного индивида представляет собой набор различающихся по длине фрагментов, соответствующих минисателлитным последовательностям его генома. Ввиду большого разнообразия этих повторов вероятность того, что в популяции найдется два человека с идентичными «ДНК-отпечатками", равна 10~5—10~8. Другими словами, характер расположения полос минисателлитных ДНК почти столь же индивидуален, как и отпечатки пальцев. Геномную дактилоскопию применяют также при установлении отцовства. Часть полос «ДНК-отпечатка» ребенка должна соответствовать полосам материнского «отпечатка», а часть — отцовского.
Если ДНК в исследуемом образце недостаточно, но она не очень сильно разрушена, можно амплифицировать небольшие участки мини-сателлитной ДНК с помощью ПЦР, а затем провести их секвенирование; этот метод более

|
| Рис. 9.7. Использование Саузерн-гибридизации для судебной экспертизы. ДНК, выделенная из крови пострадавшего, из пятна крови на рубашке подозреваемого и из его крови была обработана одной и той же рестрицирующей эндонуклеазой. «Лестница фрагментов» для ДНК, выделенной из пятна крови на рубашке, идентична таковой для ДНК пострадавшего и отличается от «лестницы фрагментов" для ДНК подозреваемого. |
194 ГЛАВА 9
чувствителен, чем определение полиморфизма длины тандемных повторов.
Использование полиморфных ДНК-маркеров
Метод «ДНК-отпечатков» может оказаться полезным и при установлении различий между растительными культурами. Один из вариантов этого метода основан на использовании полиморфных ДНК-маркеров для амплификации случайных фрагментов {random amplified polymorphic DNA, RAPD). Для этого берут произвольные праймеры длиной 9—10 нуклеотидов, не содержащие палин-дромных последовательностей и имеющие ОС-содержание 50—80%, и добавляют их по отдельности к препаратам хромосомной ДНК растений. Каждая ПЦР инициируется одним праймером, который должен быть способен связываться с обеими цепями ДНК-мишени. Нуклеотидные последовательности всех олигонуклеотидов известны, но какой из них окажется эффективным инициатором ПЦР, неясно. Если праймер гибридизуется с обеими цепями ДНК-мишени в подходящей ориентации и сайты расположены на расстоянии от 100 до 3000 п. н. друг от друга, то фланкированный ими сегмент ДНК будет амплифицирован, а
полученный фрагмент можно выделить с помощью гель-электрофореза и визуализировать окрашиванием. Число разных фрагментов ДНК, образующихся при амплификации, зависит от праймера и геномной ДНК. Для одних и тех же праймера и ДНК-мишени продукты амплификации будут каждый раз одинаковыми, а замена лишь одного нуклеотида в праймере приведет к полной смене набора получаемых фрагментов. Таким образом, используя один и тот же набор олигонуклеотидных праймеров, можно сравнивать RAPD-«ДНК-отпечатки» разных растительных культур, а следовательно, и сами культуры. Для выявления различий между двумя очень близкими сортами или культурами растений часто приходится использовать несколько произвольных праймеров с известной нуклеотидной последовательностью (рис. 9.8). Как и все другие молекулярные маркеры, RAPD можно применять для характеристики целых геномов, отдельных хромосом или генов.
По сравнению с другими методами идентификации сложных ДНК метод RAPD обладает следующими преимуществами: 1) для всех видов растений можно использовать один и тот же (универсальный) набор олигонуклеотидных праймеров; 2) не нужно создавать геномные биб-

| Рис. 9.8. Электрофорез ПЦР-амплифицированных фрагментов растительной ДНК в полиакриламидном геле с последующим окрашиванием бромистым этидием. Для амплификации фрагментов каждой из двух культур использовали три разных произвольных праймера. В случае праймеров А и В характер распределения полос в полиакриламидном геле для культур 1 и 2 совпадает, если же используется праймер Б, то положение полос различается. Таким образом, с помощью праймера Б можно выявлять различия между культурами 1 и 2. |
Молекулярная диагностика 195
лиотеки, использовать радиоактивные зонды, проводить гибридизацию, т. е. можно легко и быстро охарактеризовать большое количество образцов; 3) процесс можно автоматизировать. Кроме того, для проведения обычной ПЦР необходимо знать нуклеотидную последовательность искомого гена или его фрагмента — мишени для амплификации. В случае же RAPD амплифицируется любой участок генома, содержащий две комплементарные праймеp у последовательности, которые фланкируют сегмент ДНК длиной от 100 до 3000 п. н.
С помощью метода RAPD удалось отличить друг от друга шесть инбредных линий кукурузы и показать, что ПЦР-продукты гибридов кукурузы представляют собой сочетание ПЦР-продуктов родительских инбредных линий. RAPD-маркеры использовали также для скрининга разных штаммов грибов Leptosphaeria maculans, вызывающих заболевание «черная ножка» у крестоцветных. Было установлено различие между невирулентным (не приводящим к развитию болезни) и вирулентным (вызывающим заболевание) штаммами.
Молекулярная диагностика генетических заболеваний
Диагностика специфических наследственных заболеваний человека на генетическом уровне дает ответ на вопрос, входят ли обследуемые индивидуумы или их потомки в группу повышенного генетического риска. ДНК-анализ можно использовать для выявления носителей генов наследственных заболеваний, а также для пренатальной и пресимптоматической диагностики серьезных генетических нарушений.
Тесты на уровне ДНК позволяют безошибочно выявлять специфические мутации. Раньше для этого применялись биохимические методы, основанные на выявлении продукта анализируемого гена. ДНК-тесты не требуют экспрессии мутантного гена для его выявления, что позволяет разработать системы скрининга для всех моногенных заболеваний.
Серповидноклеточная анемия
Серповидно клеточная анемия ~ генетическое заболевание, обусловленное заменой одного нуклеотида в кодоне, который соответствует шестой аминокислоте в ß-цепи молекулы гемоглобина. У индивидов, гомозиготных по мутантному гену (S/S), эритроциты имеют необычную серповидную форму; это связано с искажением конформации молекулы гемоглобина вследствие замены в ней валина на глутаминовую кислоту. Мутантный гемоглобин не может с достаточной эффективностью переносить кислород, и у таких больных развивается тяжелая анемия с прогрессирующим поражением сердца, легких, мозга, суставов и других органов. У индивидов, гетерозиготных по данному гену (A/S) (носителей генетического заболевания), эритроциты имеют нормальную форму, и симптомы заболевания проявляются лишь в экстремальных условиях (на большой высоте над уровнем моря либо при слишком высоких или низких температурах, когда снижается снабжение организма кислородом). Если оба родителя гетерозиготны (имеют генотип A/S), то вероятность того, что их ребенок будет гомозиготным по мутантному гену (S/S) (т. е. будет болен серповидноклеточной анемией), составляет 25%. Ген серповидноклеточной анемии с высокой частотой встречается среди афроамериканцев и их потомков, а также среди латиноамериканцев. В США проводят скрининг для выявления носителей гена серповидноклеточной анемии, которые могут передать этот ген своим потомкам. Рассмотрим один из используемых для этого тестов.
Замена одного нуклеотида в ß-глобиновом гене, приводящая к серповидноклеточной анемии, сопровождается элиминацией сайта для рестрицирующей эндонуклеазы CvnI. Этот фермент узнает последовательность CCTNAGG и расщепляет молекулу ДНК между основаниями С и Т (N -любой из четырех нуклеотидов). В нормальном гене эта последовательность имеет вид CCTGAGG, а в гене серповидноклеточной анемии — ССТ-GTGG. На этом различии основывается ДНК-диагностика данного заболевания (рис. 9.9).
Используя праймеры, фланкирующие сайт Cvn1, амплифицируют с помощью ПЦР небольшое количество тестируемой ДНК (рис. 9.9, А), Амплифицированный фрагмент обрабатывают Cvnl, продукты рестрикции разделяют с помощью гель-электрофореза и окрашивают их бромистым этидием. При наличии Сvn1-сайта на электрофореграмме появляется специфический набор полос (рис. 9.9, В), отличный от такового
196 ГЛАВА 9

| Рис. 9.9, Выявление мутантного гена, ответственного за развитие серповидноклеточной анемии, А. ПЦР-амплификация участка ß-глобинового гена, содержащего сайты для эндонуклеазы CvnI, один из которых отсутствует в му-тантном гене. Б. Рестрикция полученных ПЦР- продуктов с помощью CvnI. Нормальный ген содержит три Cvn I-сайта в сегменте ДНК, фланкируемом прай-м ерам и, а мутантный — два. B. Электрофоретическое разделение фрагментов, полученных при обработке ПЦР-амплифицированной р-глобиновой ДНК с помощью Cvnl. АА — гомозиготность по нормальному β- гпоби новому гену, AS — гетерозиготност:ь, SS — гомозитотность по гену серповид-никлеточной анемии. |
в отсутствие Cvnl-сайта. Описанным способом можно без труда и достаточно быстро установить генетический статус обследуемого, не проводя при этом процедуру гибридизации,
Метод ПЦР/ЛОЗ
Не все генетические нарушения, приводящие к появлению дефектных генов, сопровождаются утратой или изменением сайтов рестрикции, поэтому для обнаружения однонуклеотидных замен применяют и другие подходы. В одном из них объединены ПЦР и метод, основанный на лигировании олигонуклеотидных зондов (ЛОЗ), ПЦР/ЛОЗ.
Дата добавления: 2015-04-16; просмотров: 297; Мы поможем в написании вашей работы!; Нарушение авторских прав |