КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Пример 2. Почему у орхидей такие разнообразные цветки?
Семейство орхидные (Orchidaceae) – второе по величине среди цветковых растений. Оно насчитывает около 30 тысяч видов, поражающих разнообразием и изысканностью своих цветков. За их красотой, однако, стоит суровая утилитарность: цветок орхидеи приспособлен для опыления строго определённым опылителем (60% видов орхидей опыляются лишь одним видом животных, главным образом насекомых). Но почему именно у орхидных (а не у лилейных, например) появлось такое разнообразие цветков? Ответ на этот вопрос дают недавние работы М. Мондрагон-Паломино с соавторами (Mondragón-Palomino & Theißen, 2008, 2009; Mondragón-Palomino, 2009), проливающие свет и на происхождение этого огромного семейства.
При всем своем разнообразии цветки орхидей имеют сходный план строения: как правило, их трудно перепутать с цветками растений из других семейств. Для подавляющего большинства орхидей характерна отчетливая двусторонняя симметрия цветка. Околоцветник всегда состоит из шести элементов, расположенных в два круга по три. Обычно листочки наружного круга околоцветника (иногда неверно называемые «чашелистиками») примерно одинаковы по форме и размерам (T1-T3 на схеме внизу), в то время как средний элемент внутреннего круга (t3, губа) как правило резко отличается от двух боковых листочков (t1 и t2). Губа играет важную роль во взаимодействии с опылителем, а потому её строение бывает особенно замысловатым.
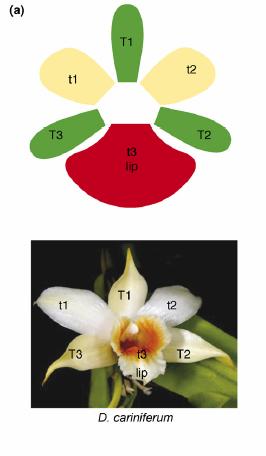
Схема строения околоцветника орхидей на примере Dendrobium cariniferum (из статьи Mondragón-Palomino & Theißen, 2008)


Слева: радиально-симметричный цветок у Spiloxene serrata, представителя семейства гипоксиевые (Hypoxidaceae) из Южной Африки. Справа: симметрия цветка орхидей Apostasia wallichii (подсемейство Apostasioideae) из Новой Гвинеи близка к радиальной (http://www.orchidspecies.com/apostwallichii.htm).




Орхидеи: ваниль (Vanilla planifolia, подсемейство Vanilloideae) из тропиков Нового Света (http://www.dragonagro.com/wholesale_orchids.htm); Венерин башмачок (Cypripedium calceolus, подсемейство Cypripedioideae), Ленинградская область; ятрышник шлемоносный (Orchis militaris, подсемейство Orchidoideae), Ленинградская область; дремлик болотный (Epipactis palustris, подсемейство Epidendroideae), Ленинградская область.
Ближайшим родственником орхидных является семейство гипоксиевые (Hypoxidaceae); к нему относятся травянистые растений с радиально-симметричными цветками. Само же семейство Orchidaceae состоит из пяти подсемейств: Apostasioideae (небольшая группа орхидей, встречающихся в Непале, Японии, Китае, Новой Гвинее и Австралии), Vanilloideae (например, ваниль), Cypripedioideae (сюда относится венерин башмачок), Orchidoideae (например, ятрышник или любка) и Epidendroideae (80% видов орхидей, в их числе дремлик, гнездовка, тайник и др.). Эти подсемейства отчетливо различаются по особенностям цветка; кроме того, их выделение подтверждено данными молекулярной филогенетики. Среди них подсемейство Apostasioideae - явно самое древнее и примитивное: венчик у его представителей почти радиально-симметричный, есть и ряд других архаичных особенностей.

Родственные связи между Hypoxidaceae и подсемействами орхидных (из статьи Mondragón-Palomino & Theißen, 2008). Филогенетическое дерево, построено путем анализа последовательностей ДНК нескольких пластидных генов (rbcL, matK, atpB), взятых у большого числа видов из всех перечисленных семейств и подсемейств. Хотя эти гены не имеют отношения к развитию цветка, полученная на их основе схема родственных связей хорошо согласуется с имеющимися представлениями об эволюции этого органа у орхидных, в частности – с тенденцией к переходу от радиальной к двусторонней симметрии околоцветника.
У всех изученных цветковых растений в регуляции образования околоцветника участвуют так называемые гены В-класса, подобные генам DEFICIENS (DEF) и GLOBOSA (GLO) львиного зева (им соответствуют APETALA3 и PISTILLATA у Arabidopsis). Мы не будем обсуждать АВС-модель и более современные концепции, объясняющие работу этих генов (подробности можно посмотреть здесь или здесь). Нам важно лишь отметить, что у многих растений имеется только один DEF-подобный и один GLO-подобный ген, которые и обеспечивают формирование у них нормальных цветков.
У орихидных, однако, дело обстоит гораздо интереснее. Как обнаружила М. Мондрагон-Паломино и её коллеги, у них есть целых четыре DEF-подобных гена, и каждый из них выполняет свою функцию. Условно говоря, гены 1 и 2 определяют, что органы, формирующиеся под их контролем, станут именно листочками околоцветника, а не листьями, тычинками или плодолистиками; ген 3 задаёт отличия элементов внутреннего круга околоцветника от наружного, а ген 4 отвечает за образование губы и ее отличия от боковых листочков. Столь сложная система регуляции и обеспечивает разнообразие морфогенетических возможностей для цветков орхидных. Мы видим, в частности, что за формирование губы у орхидей отвечает спецальный ген: не случайно некоторые морфологи трактовали эту часть околоцветника как совершенно особый орган.
Четыре DEF-подобных гена обнаружены у представителей всех четырех подсемейств орхидных (Vanilloideae, Cypripedioideae, Orchidoideae и Epidendroideae), исследованных М. Мондрагон-Паломино. У гипоксиевых же найдены только два таких гена, причем один из них близок по структуре к генам 1 и 2 у орхидей, а другой – к их генам 3 и 4. Эти данные показывают, что у предков орхидных произошло два цикла удвоения DEF-подобных генов с последующим «разделением труда» между получившимися копиями. Вероятно, такое удвоение и стало той ключевой инновацией, которая предопределила нынешнее разнообразие и эволюционный успех орхидных.
Четыре DEF-подобных гена орхидных сравнительно сильно различаются между собой по нуклеотидным последовательностям. Это неудивительно – ведь их функции различны. Важно другое: если методами молекулярной филогенетики проанализировать последовательности любого из этих генов (скажем, 1 или 3), взятые у представителей различных подсемейств орхидных, то получатся практически одинаковые филогенетические деревья, совпадающие с деревом на рисунке, приведенном выше.
Таким образом, М. Мондрагон-Паломино и её соавторы обосновали биологически осмысленную модель происхождения и эволюции крупной и своеобразной группы растений. Эта модель демонстрирует преемственность между DEF-подобными генами гипоксиевых и орхидных, хорошо согласуется с данными как сравнительной анатомии цветка, так и с результатами молекулярно-филогенетического анализа. Мы видим, что убедительный сценарий крупного макроэволюционного события – каковым, безусловно, было появление орхидных – совершенно не нуждается в привлечении каких-либо сверхестественных факторов.
Впрочем, мы пока ничего не знаем про гены, регулирующие формирование околоцветника у Apostasioideae, самой примитивной группы орхидных. Очень интересно, сколько DEF-подобных генов окажется у них. На основе имеющегося эволюционного сценария можно предположить, что этих генов окажется либо два (как у гипоксиевых), либо четыре (как у остальных орхидных), либо три (ведь апостасиевые вполне могут рассматриваться как переходная группа между двумя семействами), но никак не один и не больше четырех. Время делать ставки!
Литература:
Mondragón-Palomino M., Theißen G. 2008. MADS about the evolution of orchid flowers. Trends in Plant Science 13(2): 51-59.
Mondragón-Palomino M., Theißen G. 2009. Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes. Annals of Botany, advanced online access, Jan 13 2009.
Mondragón-Palomino M., L. Hiese, A. Härter, M.A. Koch, and G. Theiβen. 2009. Positive selection and ancient duplications in the evolution of class B floral homeotic genes of orchids and grasses. BMC Evolutionary Biology 9:80
Дата добавления: 2015-04-16; просмотров: 366; Мы поможем в написании вашей работы!; Нарушение авторских прав |