КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ЭУКАРИОТНЫХ ОРГАНИЗМОВ
В.В.Малахов
ОСНОВНЫЕ ЭТАПЫ ЭВОЛЮЦИИ
ЭУКАРИОТНЫХ ОРГАНИЗМОВ
©2003г.
Московский государственный университет
Поступила в редакцию 10.02.2003 г.
Принята к печати 02.04.2003 г.
ПАЛЕОНТОЛОГИЧЕСКИЙ ЖУРНАЛ, 2003, № 6, с. 25-32
Обсуждаются основные этапы эволюции эукариот. Принципиальным моментом в происхождении эукариот было появление актиново-миозиновой системы. Благодаря наличию этой системы предки эукариот приобрели способность к формированию псевдоподий и хищному питанию путем фагоцитоза. Ядерная оболочка появилась для защиты генетического материала в условиях высокой подвижности цитоплазмы эукариот. Митохондрии и пластиды происходят от симбионтов. Пластиды зеленых и красных водорослей - потомки различных прокариотных фотосинтезирующих бактерий. Пластиды криптомонад, хризомонад, бурых водорослей, динофлагеллят и эвгленовых происходят от различных эукариотических организмов. Предки высших растений - многоклеточные зеленые водоросли, которые обитали в мелких пересыхающих водоемах на суше или на влажном грунте. В этих условиях размножение зооспорами неэффективно. Спорофит высших растений первоначально был органом распространения воздушных спор. Отсутствие жгутиковых стадий у настоящих грибов позволяет принять их происхождение в наземных экосистемах. Предполагается, что многоклеточные животные происходят от морских колониальных воротничковых жгутиконосцев. Обсуждается гипотеза происхождения билатерий от кишечнополостных, ползавших на оральной поверхности.
В настоящее время считается твердо установленным, что большую часть своего существования, вплоть до так называемой неопротерозойской революции (около 1 млрд. лет назад), биосфера была прокариотной (Заварзин, 2001). В такой биосфере и продуценты, и редуценты были представлены только прокариотными организмами. Биомасса, созданная автотрофными бактериями, подвергалась разложению преимущественно под действием абиотических физических и химических факторов внешней среды. Нет сомнения, что значительную роль в разложении биомассы, созданной прокариотными автотрофами, играли и гетеротрофные бактерии, однако их возможности были ограничены в силу особенностей организации клеток прокариот.
Как известно, прокариоты принципиально не способны к хищничеству путем заглатывания своих жертв. Хищничество у бактерий изредка встречается, но носит весьма своеобразный характер (рис. 1, а). Так, например, бактерия Bdellovibrio - это маленький вибрион, который проникает через клеточную стенку жертвы (значительно более крупной, чем напавший на нее хищник) и размножается в промежутке между ее клеточной стенкой и мембраной (Stolp, 1979). Как своеобразное хищничество можно рассматривать и нападение фагов на бактерий. Но ведь и в этом случае фаг только впрыскивает в цитоплазму жертвы свою нуклеиновую кислоту, а белковая оболочка фага остается снаружи (рис. 1, б). Прокариоты не способны даже к поглощению пузырьков жидкости, т.е. к пиноцитозу.

Не обладая способностью заглатывать свои жертвы целиком или по кусочкам, гетеротрофные прокариоты могли использовать только эк-зоферментацию субстрата с последующим поглощением низкомолекулярных органических веществ. Все это, как можно предполагать, обусловливало низкую скорость разложения биомассы, созданной автотрофными прокариотами. Огромные массы органического углерода захоранивались в осадке и на долгое время выводились из биотического круговорота.
Почему же прокариоты не обладают способностью к фагоцитозу и пиноцитозу? Дело в том, что у прокариот отсутствует актиново-миозиновая система. Актин и миозин - универсальные сократительные белки клеток эукариот. Они обеспечивают амебоидную активность, движение органелл внутри клетки (так называемый "циклоз"), мышечные сокращения и другие формы клеточной подвижности эукариот (Каппучинелли, 1982). Актиново-миозиновая система обеспечивает способность образовывать псевдоподии, захватывать ими жертву и формировать пищеварительные вакуоли (рис. 1, в). Даже вирусы проникают в клетку эукариот путем провокации эндоцитоза - активного захвата их клеткой (рис. 1, г). Таким образом, можно утверждать, что становление эукариот было связано с появлением актиново-миозиновой системы, которая позволила питаться путем фагоцитоза, активно захватывая крупные пищевые частицы. Первые эукариоты - это фаготрофные организмы, способные к амебоидному движению (Мирабдуллаев, 1989а, 1992).
Появление таких эукариотных организмов необычайно ускорило биотический круговорот, поскольку они стали потребителями бактериальной биомассы. Переваривая клетки бактерий, фаготрофные эукариоты быстро возвращали в биотический круговорот элементы, которые до этого могли вернуться в него только путем медленного разложения. Можно предполагать, что появление эукариот означало резкое уменьшение "бактериальных ископаемых", т.е. отложений органических и неорганических веществ, возникших за счет деятельности бактерий.
Способ питания эукариот путем захвата пищевых частиц означал, что хищник был крупнее жертвы. Действительно, линейные размеры мелких почвенных амеб или жгутиконосцев, питающихся бактериями, приблизительно в 10 раз больше размеров бактерий. Таким образом, объем цитоплазмы эукариот приблизительно в 1000 раз больше, чем у прокариот. Такой большой объем цитоплазмы требовал и большого числа копий генов, чтобы снабжать увеличенную цитоплазму продуктами транскрипции. Один из способов решения этой задачи - умножение числа генофоров. Полиплоидия - один из путей увеличения числа копий генов у эукариот, но этот путь известен и для прокариот (так называемые "полиплоидные бактерии"- см. Пешков, 1966). Вероятно, и предки эукариот с большим объемом цитоплазмы пошли по пути мультипликации генофора (рис. 2). Множественные генофоры (первоначально одинаковые) стали зачатками хромосом, в которых постепенно накапливались различия.
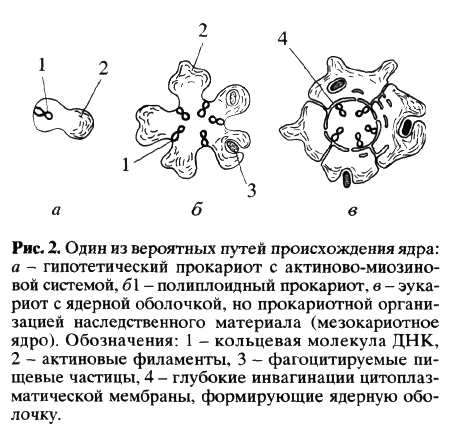
При амебоидном движении и питании путем фагоцитоза цитоплазма клетки (особенно периферическая) оказывается очень подвижной. Генофоры, прикрепленные к поверхностной мембране клетки, оказывались в зоне сильных токов цитоплазмы. Вот почему появился защищенный мембранами участок в центральной цитоплазме, в котором хранились генофоры. Этот процесс мог происходить по-разному, но один из возможных путей - глубокие впячивания участков цитоплазматической мембраны с прикрепленными к ним генофорами (ведь ядерная оболочка - это часть эндоплазматического ретикулюма эукариотной клетки, который может быть связан с внешней средой). Первичные эукариоты, таким образом, имели ядро, ограниченное двойной ядерной оболочкой - производным эндоплазматического ретикулюма, но имели еще кольцевое строение генофоров и были лишены гистонов (рис. 2, el). Как это ни удивительно, подобное строение ядра действительно сохранилось у некоторых современных эукариот, например у динофлагеллят. Как известно, у этих протистов ядро окружено двойной ядерной мембраной, но хромосомы содержат кольцевые (хотя и политенные) молекулы ДНК, практически лишенные гистонов (Райков, 1978).
Способность к фаготрофному питанию предопределила возможность появления у эукариот клеточных эндосимбионтов. Не обладая способностью к заглатыванию жертв, прокариоты не могли приобрести и клеточных эндосимбионтов. Для эукариот, наоборот, включение в качестве эндосимбионтов прокариотных и эукариотных организмов весьма обычно и происходило многократно и на различных уровнях организации. В настоящее время старая идея (Mereschkowsky, 1905; Фаминицын, 1907; Мережковский, 1909) о симбиотическом происхождении некоторых органелл эукариотной клетки (митохондрий и пластид) стала одной из парадигм биологии (Margulis, 1970,1981,1992; Тахтаджян, 1973; Мирабдуллаев, 1985, 19896; Margulis et al., 1999).
Согласно одной из распространенных гипотез, в результате установления симбиоза с оксифильными бактериями - предками митохондрий - эукариоты приобрели возможность существовать в обогащенной кислородом атмосфере позднего протерозоя, а приобретение пластид привело к появлению в биосфере разнообразных эукариотных автотрофов.
Эукариотные организмы с растительным типом обмена (водоросли, мхи и сосудистые растения) появились в результате симбиоза хищных протестов с различными фотосинтезирующими организмами (Raven, 1970; Whatley et al., 1979; Мирабдуллаев, 1985). Фотосинтезирующие прокариоты - цианобактерии, вероятно, дали начало пластидам красных водорослей Rhodophyta. Пластиды у них окружены только двумя мембранами (внешняя - мембрана вакуоли хозяина и внутренняя - мембрана симбионта), содержат типичные для цианобактерии хлорофилл а и фикобилины, а тилакоиды не образуют ламелл (рис. 3, а, 6). Глаукофитовые жгутиконосцы (Glaucophyta) также приобрели цианобактерии в качестве эндосимбионтов, притом совершенно независимо от красных водорослей. У Glaucophyta эндосимбиотические цианеллы сохраняют еще муреиновую оболочку, характерную для свободноживущих цианобактерии (Herdman, Stanier, 1977).
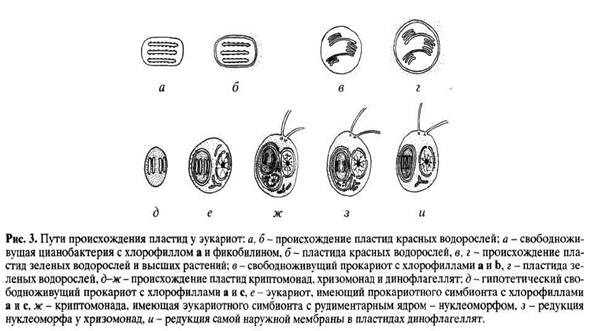
Зеленые водоросли, мхи и сосудистые растения также имеют пластиды, организация которых говорит об их прокариотном происхождении. Они окружены двумя мембранами, но содержат другой набор пигментов, а именно: хлорофиллы а и b. Это дает основание предполагать, что пластиды зеленых водорослей не родственны цианобактериям, а произошли от другой группы фотосинтезирующих оксибактерий, обладающих хлорофиллами а и b (рис. 3, в, г). И действительно, в современной биосфере найдены прокариотные организмы, обладающие хлорофиллами а и b и тилакоидами, собранными в ламеллы. Это представители особой группы сравнительно недавно обнаруженных оксифотобактерий Prochlorophytes (Lewin, 1976, 1977, 1984; Whatley, 1977; Burger-Wiersma et al., 1986; Chisholm et al., 1988). В современной биосфере они занимают довольно скромное место, хотя разнообразие жизненных форм этих бактерий (это и обитатели туники сложных асцидий, и свободноживущие планктонные формы, и нитчатые бактерии) говорит о том, что прохлорофиты в прошлом могли играть роль, не менее важную, чем цианобактерии. Возможно, что прохлорофиты могли участвовать в образовании строматолитов (Reimer et al., 1979).
Современные прохлорофиты не являются предками пластид зеленых водорослей (об этом говорят, в частности, данные анализа нуклеотидной последовательности генов рибосомальных РНК (Seewaldt, Stackebrandt, 1982); скорее всего, и те и другие имели отдаленного общего предка.
Пластиды золотистых водорослей (Chrysophyta), желто-зеленых водорослей (Xanthophyta), бурых и диатомовых водорослей устроены морфологически сложнее, чем пластиды красных и зеленых водорослей. Они окружены четырьмя мембранами и содержат другой набор хлорофиллов, а именно: a, cl и с2. Происхождение таких пластид было непонятно до тех пор, пока не были исследованы пластиды криптофитовых водорослей (Cryptophyta). Последние имеют тот же набор хлорофиллов и четырехмембранную оболочку. Своеобразие пластид криптофитовых состоит в том, что в пространстве между второй и третьей мембранами у них обнаружено маленькое эукариотное ядро (так называемый "нуклеоморф") и цистерны эндоплазматического ретикулюма (Gillot, Gibbs, 1980; Cavalier-Smith, 1981; Gibbs, 1981). Это позволяет предполагать, что предками пластид криптофитовых были эукариотные организмы, которые сами в свое время приобрели прокариотных симбионтов. Таким образом, клетка криптофитовой водоросли является сложной конструкцией: она содержит эукариотного симбионта, внутри которого находится прокариотный симбионт. Наружная мембрана пластиды это мембрана вакуоли хозяина, вторая мембрана это поверхностная мембрана эукариотного симбионта, третья мембрана это мембрана вакуоли эукариотного симбионта, а четвертая это мембрана прокариотного симбионта. Следовательно, сначала какой-то хищный протист приобрел прокариотного симбионта - фотосинтезирующую бактерию с хлорофиллами a, cl и с2, а потом сам этот протист стал симбионтом другого эукариотного жгутиконосца (рис. 3, д-ж).
Пластиды Xanthophyta, Phaeophyta и Bacillariophyta утратили нуклеоморф, хотя и сохранили сложную четырехмембранную оболочку. Редукция нуклеоморфа означает увеличение интеграции симбиотической системы, в процессе которой большинство генов симбионта переходит в ядро хозяина. Следующий шаг - постепенное уменьшение числа мембран симбионта. Так, у динофитовых водорослей (Dinophyta) пластиды (содержащие хлорофиллы а и с2) окружены тремя мембранами, что можно рассматривать как результат редукции одной из мембран четырехмембранной оболочки. Скорее всего, редуцирована самая наружная мембрана, т.е. та, что соответствует мембране вакуоли хозяина (рис. 3, д-ж). В пользу этого говорит то, что в отличие от обычных четырехмембранных пластид, наружная мембрана пластид динофитовых никогда не несет рибосом и не связана с ядерной оболочкой хозяина (Dodge, 1979; Cavalier-Smith, 1981). Заметим, что вплоть до настоящего времени остается проблематичным, существуют ли в современной биосфере свободноживущие прокариоты, близкие к предкам пластид золотистых и бурых водорослей. Возможно, все свободноживущие представители этой группы давно уже вымерли либо сильно изменились. Возможно, некоторым сходством с хлоропластами золотистых и бурых водорослей обладает анаэробная азотофиксирующая бактерия Heliobacterium chlorum (см. Рейвн и др., 1990).
Любопытно, что аналогичные эволюционные процессы идут и в ряду водорослей с зелеными пластидами, содержащими хлорофиллы а и b. Так, у амебоидного организма Clorarachnion reptans имеются зеленые пластиды, окруженные четырьмя мембранами и содержащие нуклеоморф (Hibberd, Norris, 1984). Эвгленовые водоросли (Euglenophyta) имеют зеленые пластиды с тремя мембранами, что также можно рассматривать как результат редукции самой наружной мембраны, соответствующей стенке вакуоли хозяина (Gibbs, 1978).
Эукариотные водоросли, появившиеся, как можно думать, около 1 млрд. лет назад, существенно потеснили прокариотные фотосинтезирующие организмы и с того времени стали главными (но все же не единственными!) производителями органического вещества в эуфотической зоне Мирового океана. Во всех основных эволюционных линиях эукариотных водорослей возникли многоклеточные формы. Золотистые водоросли (Chrysophyta) вероятно дали начало бурым водорослям (Phaeophyta), талломы которых достигают 60 м в длину. Сложнорасчлененный многоклеточный таллом характерен для красных водорослей. В пределах зеленых водорослей Chlorophyta многоклеточные формы возникали независимо несколько раз (Chlorophycea, Charophycea, Ulvophycea и др.). Многоклеточные бурые водоросли, многоклеточные красные водоросли и многоклеточные зеленые водоросли (от которых берут начало высшие растения) не родственны друг другу. Вот почему невозможно создать монофилетическое царство многоклеточных растений Metaphyta. Приходится иметь дело с царством красных водорослей Rhodobiontae, царством золотисто-бурых водорослей Chromobionthae, царством зеленых растений Viridiplantae.
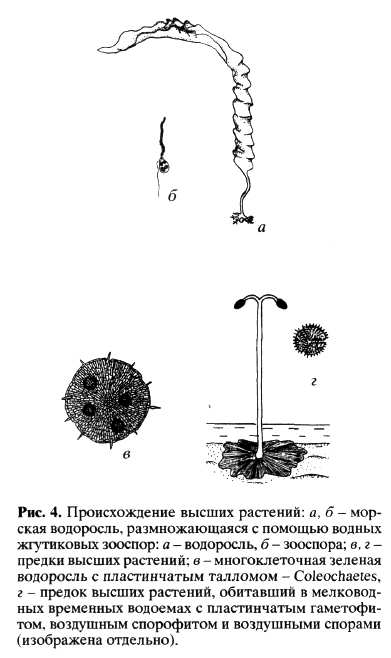
Сосудистые растения и мхи это несомненные потомки зеленых водорослей Chlorophyta. Происхождение высших растений связано с освоением суши, и ему предшествовало заселение внутриконтинентальных водоемов разнообразными зелеными водорослями. Следует учесть, что гидрография суши в докембрии и раннем палеозое существенно отличалась от современной (Пономаренко, 1993; Еськов, 2000). Отсутствие растительного покрова приводило к быстрому стоку всех осадков, поэтому в это время, вероятно, не было постоянно существующих рек и озер. Биотопами для водорослей были мелководные временные водоемы или влажный грунт на месте временного водоема. В этом случае размножение обычными для водорослей жгутиковыми зооспорами было неэффективно, поскольку возможности расселения ограничены только размерами самого временного водоема. Для заселения пересыхающих биотопов необходимо иметь споры, переносимые по воздуху. Спорофит высших растений, по-видимому, возник первоначально только как орган для распространения воздушных спор (рис. 4). При этом гаметофит мог быть чисто водным или обитать на влажном грунте. Пластинчатая форма гаметофита примитивных печеночных мхов и папоротников, вероятно, рекапитулирует организацию таллома предков высших растений, обитавших на дне очень мелких временных водоемов или на влажном грунте. Кстати, пластинчатый таллом характерен для колеохетовых водорослей (Соleochaetales - см. рис. 4), которые по совокупности признаков ближе всего стоят к высшим растениям (Graham, 1984, 1985).
Фаготрофные протисты демонстрируют различные способы выхода за пределы одноклеточности: многоядерные жгутиковые (например, Opalinata), полиплоидные простейшие (например, Radiolaria), ресничные простейшие (Ciliophora), клеточные агломераты (Acrasidea), плазмодии (Mycetozoa) и др. (Иванов, 1968; Шульман, 1974). Причины этого понятны: крупные формы могут питаться более крупной добычей, а сами при этом оказываются недоступны для мелких хищников. К многоклеточным животным (Metazoa) привел, по-видимому, только один путь - создание подвижных колоний одноядерных жгутиковых простейших.
В отличие от растительных организмов, где многоклеточность возникала несколько раз, происхождение многоклеточных животных, по-видимому, связано только с одной группой фаготрофных протистов, а именно - с воротничковыми жгутиконосцами Choanoflagellata. Таким образом, царство многоклеточных животных с высокой степенью вероятности - монофилетическое, а сами Metazoa являются настоящим таксоном.
Воротничковые жгутиконосцы обладают единственным жгутом (что характерно и для многоклеточных животных), вокруг которого располагается венчик тентакул-микроворсинок (рис. 5, а). Последние представляют собой нитевидные выросты цитоплазмы (имеющие внутренний скелет из микрофиламентов) и отличаются как от псевдоподий, так и от аксоподий других простейших. Микроворсинки образуют ловчий воротничок, с помощью которого захватываются бактерии, принесенные токами воды, созданными биением жгута (Жуков, Карпов, 1985). Такую же функцию выполняют микроворсинки воротничковых клеток (хоаноцитов) низших многоклеточных - губок. Зачаточный воротничок характерен для жгутиковых эктодермальных клеток кишечнополостных, клеток жгутиковой целомической выстилки вторичноротых, а также сенсорных клеток в различных группах Metazoa. В других случаях микроворсинки утрачивают правильное круговое расположение, но остаются почти обязательным элементом строения апикальной поверхности эпителиальных клеток многоклеточных животных.

Первичные Metazoa, вероятно, были однослойными шарообразными организмами, медленно плавающими в толще воды (рис. 5, б). Организацию таких предковых форм рекапитулируют бластулообразные личинки многоклеточных животных (рис. 5, в). Однослойные докембрийские Metazoa, перейдя к прикрепленному образу жизни, дали большое разнообразие форм в позднем докембрии (Бурзин, 1999). Возможно, некоторые трудно интерпретируемые организмы венда представляют собой результат морфологического усложнения тонкостенных потомков бластулообразных предков Metazoa. От них же, по-видимому, происходят губки и губкообразные организмы (археоциаты) фанерозоя. В отличие от тонкостенных организмов докембрия у губок усиленному развитию подвергся мезохил - промежуточное вещество бластоцеля, в котором возникли скелетные элементы и который постепенно заселился клетками различных типов.
По типу захвата пищевых частиц губки (как, вероятно, и тонкостенные организмы венда) не отличаются от одноклеточных протестов: пищевые частицы (бактерии) захватываются каждой воротничковой клеткой индивидуально и перевариваются в пищеварительных вакуолях. Поэтому и вся эволюция примитивных многоклеточных оказывается направленной на увеличение поверхности слоя воротничковых клеток, через который происходит захват пищевых частиц. У губок - это формирование многочисленных жгутиковых камер в толще мезохила, общая площадь которых превосходит наружную поверхность во много раз, а у тонкостенных вендских форм - формирование складок и камер, выстланных питающим эпителием.
Настоящие многоклеточные (Eumetazoa) - это формы, имеющие орган захвата пищи - рот, орган переваривания пищи - кишечную полость, нервную систему и мускулатуру. Рот, кишечник, нервная система и мускулатура появляются в эволюции как бы "одновременно". Не существует животных со ртом и кишечником, но без мускулатуры и нервной системы или организмов с нервной системой, но без мускулатуры, рта и кишечника и т.п. Вероятно, это означает, что само появление кишечной полости было связано с питанием относительно крупной добычей, для овладения которой требовались координированные мышечные сокращения и нервная система. Первыми организмами, обладающими нервной системой, мускулатурой, ртом и кишечником, были вендские кишечнополостные.
Вендские Coelenterata это кишечнополостные в широком смысле, т.е. двуслойные организмы с кишечником, связанным с внешней средой только ротовым отверстием. Среди них были и радиально-симметричные и билатерально-симметричные формы. Нет доказательств, что вендские кишечнополостные обладали специализированными стрекательными клетками или гребными пластинками, т.е. нет оснований относить их к типам Cnidaria или Ctenophora. Скорее всего, стрекающие кишечнополостные и гребневики появились уже в фанерозое, хотя их предками были вендские кишечнополостные.
Как известно, в отложениях венда имеются и первые билатерально-симметричные формы (Dickinsonia, Vendia, Onega и др., см. Соколов, 1976,1997; Федонкин, 1981,1983,1984). Таким образом, происхождение первых билатерально-симметричных форм следует отнести еще к позднему докембрию (приблизительно 600 млн. лет назад). Морфология вендских Bilateria свидетельствует о том, что первые билатерально-симметричные животные были метамерными организмами. Эта метамерия есть результат трансформации цикломерии кишечнополостных: цикломеры превратились в сегменты. Экологической предпосылкой появления Bilateria был переход радиально-симметричных пелагических кишечнополостных к ползанию по субстрату на оральной поверхности (рис. 6, а, б). При таком образе жизни оральная сторона становится брюшной, а аборальная - спинной, появляется передний конец и новая переднезадняя ось, которая одновременно является и осью метамерии. Цикломеры радиально-симметричного предка становятся метамерами билатерально-симметричного организма. Гипотезы, предполагающие такой путь становления Bilateria, появились более ста лет назад (Sedgwick, 1884; Beneden, 1891; Masterman, 1898), но только открытие вендской биоты позволило найти для них палеонтологические доказательства.
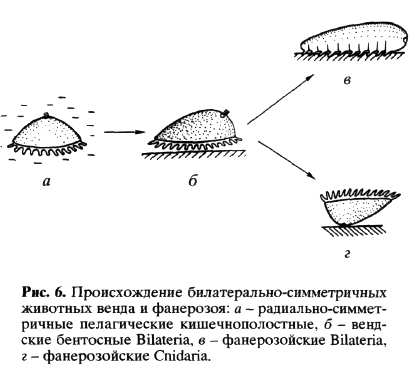
На ранних этапах эволюции Bilateria еще не имели сквозного кишечника (вероятно, сохранялся длинный щелевидный рот), а метамерно расположенные карманы гастральной полости были связаны с центральной частью пищеварительного тракта. Это означает, что вендские Bilateria еще не были трехслойными формами, т.е. оставались на уровне кишечнополостных. На этой стадии от вендских Bilateria отделились предки фанерозойских Cnidaria. Как известно, в строении и развитии кораллов (примитивной группы стрекающих, не обладающих метагенезом) отчетливо проявляются следы предковой билатеральной симметрии, которой обладали медленно ползающие на оральной поверхности вендские Bilateria. Предковые формы Cnidaria прикрепились аборальной поверхностью к субстрату, в связи с чем утратили аборальный нервный центр (рис. 6, г). Сидячий образ жизни привел к формированию радиальной симметрии Cnidaria, особенно ясно выраженной у сцифоидных и гидроидных. Каким бы парадоксальным это ни показалось, но радиальная симметрия фанерозойских Cnidaria, скорее всего, возникла вторично, на базе слабо оформленной билатеральной симметрии вендских Bilateria (рис. 6, б). Вот почему порядок симметрии, преобладающий у фанерозойских Cnidaria (4-лучевая симметрия), оказался иным, чем у вендских радиально-симметричных форм, у которых доминирует 3-лучевая симметрия (Федонкин, 1981, 1984).
У фанерозойских Bilateria (рис. 6, в) метамерные карманы гастральной полости отделились от центральной части кишечника и дали начало метамерному целому. Щелевидный рот сомкнулся в средней части, отверстия на переднем и заднем конце стали ртом и анусом. Аборальный нервный центр дал начало надглоточному (церебральному) ганглию, а околоротовое нервное сплетение туловищной нервной системе Bilateria. Все эти преобразования давно обсуждались морфологами (Goette, 1884; Lang, 1884; Беклемишев, 1944, 1964; Remane, 1950; Siewing, 1969, 1976), опиравшимися на данные эмбрионального развития и сравнительной анатомии. Открытие вендских билатерально-симметричных организмов не оставляет другой альтернативы, кроме признания того, что предки Bilateria были метамерными целомическими организмами (рис. 6).
Высшие растения и многоклеточные животные - две важнейшие экологические группы (продуценты и консументы), во многом определившие своеобразие биологического круговорота в фанерозое. Не менее важную роль в фанерозойской биосфере играет третья группа многоклеточных эукариот - грибы (Eufungi), которые являются отдельной жизненной формой среди современных многоклеточных эукариот. Они осуществляют экзоферментацию субстрата и осмотрофное питание низкомолекулярными органическими веществами. Экологической предпосылкой происхождения грибов было появление богатых органическим веществом биотопов на суше. "Сухопутная" природа грибов следует из полного отсутствия у них жгутиковых стадий. У них нет ни жгутиковых гамет, ни жгутиковых зооспор, характерных для водных простейших. Половой процесс и расселение спорами у грибов не зависит от наличия капельно-жидкой воды. Эта относительная (для роста и развития грибы нуждаются в высокой влажности воздуха) автономия от водной среды позволила грибам широко заселить почву и стать важнейшими деструкторами органического вещества.
Среди современных протистов трудно найти формы, близкие к предкам грибов. Отдаленными родственниками грибов обычно считаются хит-ридиевые (Chytridiales), но их близость к грибам отнюдь не бесспорна. Как группа, приспособленная прежде всего к наземным биотопам, грибы, вероятно, не могли появиться раньше, чем появились такие мощные сухопутные продуценты органического вещества, как высшие растения. Таким образом, вполне возможно, что происхождение грибов следует отнести к ордовику-силуру, хотя расцвет этой группы организмов, вероятно, начался еще позже.
СПИСОК ЛИТЕРАТУРЫ
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М.: Сов. наука, 1944. 492 с.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. М.: Наука, 1964. Т. 1.432 с. Т. 2.446 с.
Бурзин М.Б. Загадочный мир эдиакарских организмов // Наука в России. 1999. № 2. С. 22-28.
Есъков К.Ю. История Земли и жизни на ней. М.: МИ-РОС-МАИК, 2000. 352 с.
Жуков Б.Ф., Карпов С.А. Пресноводные воротничковые жгутиконосцы. Л.: Наука, 1985. 120 с.
Заварзин Г.А. Становление биосферы // Вестн. РАН. 2001. Т. 71. № 11. С. 988-1001.
Иванов А.В. Происхождение многоклеточных животных. Филогенетические очерки. Л.: Наука, 1968. 287 с.
Каппучинелли П. Подвижность живых клеток. М.: Мир, 1982. 126 с.
Мережковский К.С. Теория двух плазм как основа симбиогенезиса, нового учения о происхождении организмов. Казань, 1909. 102 с.
Мирабдуллаев ИМ. Эволюция пластид и происхождение цианобактерий // Журн. общ. биол. 1985. Т. 46. С. 483-490.
Мирабдуллаев ИМ. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341-356.
Мирабдуллаев ИМ. Рибосомы, кристы и филогения низших эукариот // Изв. АН СССР. Сер. биол. 19896. № 5. С. 689-700.
Мирабдуллаев ИМ. К вопросу о содержании понятия "животное"//Журн. общ. биол. 1992. Т. 53. С. 662-671.
Пешков М.А. Сравнительная цитология синезеленых водорослей, бактерий и актиномицетов. М.: Наука, 1966. 246 с.
Пономаренко А.Г. Основные события в эволюции биосферы // Проблемы доантропогенной эволюции биосферы. М.: Наука, 1993. С. 15-25.
Райков И.Б. Ядро простейших. Морфология и эволюция. М.: Наука, 1978. 328 с.
Рейвн П., Эверт Р., Айкхорн С. Современная ботаника. М.: Мир, 1990. Т. 1. 348 с.
Соколов Б.С. Метазоа докембрия и вендо-кембрийский рубеж // Палеонтол. журн. 1976. № 1. С. 3-18.
Соколов Б.С. Очерки становления венда. М.: КМК Лтд, 1997. 156 с.
Тахтаджян АЛ. Четыре царства органического мира // Природа. 1973. № 2. С. 22-32.
Фаминицын А.С. О роли симбиоза в эволюции организмов // Тр. СПб общ-ва естествоиспыт. 1907. Т. 38. С. 141-170.
Федонкин М.А. Беломорская биота венда (докембрийская бесскелетная фауна севера Русской платформы) // Тр. Геол. ин-та АН СССР. 1981. Вып. 342. 100 с.
Федонкин М.А. Органический мир венда // Итоги науки и техники. Стратигр. и палеонтол. 1983. Т. 12. 128с.
Федонкин М.А. Проморфология вендских Radialia // Стратиграфия и палеонтология древнейшего фанеро-зоя. М.: Наука, 1984. С. 30-57.
Шульман С.С. Проблема происхождения Metazoa // Тр. Зоол. ин-та АН СССР. 1974. Т. 53. С. 47-82.
Beneden E. van. Recherches sur la developpement des Arachnactis. Contribution a la morphologic des Ceri-anthides //Arch. Biol. Paris. 1891. T. 11. P. 115-147.
Burger-Wiersma T., Veenhuils M., Korthals HJ. etal.A new prokaryote containing chlorophylls a and b // Nature. 1986. V. 320. P. 262-264.
Cavalier-Smith T. The origin and early evolution of the eukaryotic cell // Symp. Soc. Gener. Microb. 1981. V. 32. P. 34-83.
Chisholm S.W., Olson RJ., Zettler ЕЯ., Waterbury J.B. A novel free-living prochlorophyte abundant in the oceanic euphotic zone // Nature. 1988. V. 334. P. 340-343.
Dodge J.D. The phytoflagellates: fine structure and phylog-eny // Biochemistry and Physiology of Protozoa. N.Y. etc.: Acad. Press, 1979. Vol. 1. P. 7-57.
Gibbs S. The chloroplast of Euglena have evolved from endosymbiotic green algae // Canad. J. Botany. 1978. V. 56. P. 2883-2889.
Gibbs S.P. The chloroplast of some algal groups may evolved from endosymbiotic eukaryotic algae // Ann. N.Y. Acad. Sci. 1981. V. 361. P. 193-207.
Gillot M., Gibbs S. The cryptomonad nucleomorph: its ultra-structure and evolutionary significance // J. Phycol. 1980. V. 16. P. 558-568.
Goette A. Abhandlungen zur Entwicklungsgeschichte der Tiere. Leipzig, 1884. H. 2. 215 S.
Graham L.E. Coleochaete and the origin of land plants // Amer. J. Botany. 1984. V. 71. P. 603-608.
Graham L.E. The origin of the life cycle of land plants // Amer. Sci. 1985. V. 73. P. 178-186.
Herdman M., Stonier R. The cyanelle: chloroplasts or endosymbiotic procaryote? // FEMS Microbiol. Lett. 1977. V. 1. P. 7-12.
Hibberd DJ., Morris R.E. Cytology and ultrastructure of Chlorarachnion reptans (Chlorarachniophyta divisio nova, Chlorarachniophycea classis nova) // J. Phytol. 1984. V. 20. P. 310-330.
Lang A. Die Polycladen (Seeplanarien) // Fauna und Flora Golfes von Neapel. 1884. 11 Monogr. 688 S.
Lewin RA. Prochlorophyta as a proposed new division of algae // Nature. 1976. V. 261. P. 697-698.
Lewin RA. Prochloron, type genus of Prochlorophyta // Phycologia. 1977. V. 16. P. 217-218.
Lewin RA. Prochloron: a status report // Phycologia. 1984. V. 23. P. 203-208.
Margulis L. Origin of eukaryotic cells. New Haven: Yale Univ. Press, 1970. 350 p.
Margulis L. Symbiosis in cell evolution. Life and its environment on the early Earth. San Francisco: Freeman and Co, 1981.415р.
Margulis L. Biodiversity: molecular biological domain, symbiosis and kingdom origin//Biosystems. 1992. V. 27. P. 39-51.
Margulis L., Dolan M., Schwartz V.M. Diversity of life: the five kingdoms. N.Y.: Jones and Bartlett, 1999. 256 p.
Masterman A.T. On the theory of archimeric segmentation and its bearing upon the phyletic classification the Coeloma-ta // Proc. Roy. Soc. Edinburg. 1898. V. 22. P. 270-310.
Mereschkowsky C. Uber Natur und Ursprung der Chromatophoren im Pflanzenreiche // Biol. Centralbl. 1905. B. 25, S. 593-604.
Raven P.H. A multiple origin for plastids and mitochondria // Science. 1970. V. 169. P. 641-646.
Reimer Т.О., Barghoorn E.S., Margulis L. Primary productivity in an early archean microbial ecosystems // Precambrian Res. 1979. V. 9. P. 93-104.
Remane A. Die Entstehung der Metamerie der Wirbellosen // Zool. Anz. 1950. Suppl. 14. S. 16-23. Sedgwick A. On the nature of metameric segmentation and some other morphological questions // Quart. J. Microsc. Sci. 1884. V. 24. P. 43-83.
Seewaldt E., Stackebrandt E. Partial sequence of 16S ribosomal RNA and the phytogeny of Prochloron // Nature. 1982. V. 295. P. 618-620.
Siewing R. Lehrbuch der vergleichenden Entwicklungsgeschichte der Tiere. Hamburg: Parey, 1969. 531 S. Siewing R. Probleme und neuere Erkenntnise in der Grossystematik der Wirbellosen // Verh. Deutsch. Zool. Ges. 1976. S. 59-83.
Stolp H. Interaction between Bdellovibrio and its host cell // Proc. Roy. Soc. London B. 1979. V. 204. P. 211-217.
Whatley J.M. The fine structure of Prochloron // New phy-cologist. 1977. V. 79. P. 243-257.
Whatley J.M., John P., Whatley F. From extracellular to intracellular: the establishment of mitochondria and chloroplast//Proc. Roy. Soc. London B. 1979. V. 204. P. 165-187.
Дата добавления: 2015-04-16; просмотров: 286; Мы поможем в написании вашей работы!; Нарушение авторских прав |
| <== предыдущая лекция | | | следующая лекция ==> |
| Диагностика профессиональной подготовленности к педагогической деятельности начинающего преподавателя | | | Цели и задачи промышленного маркетинга |