КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Происхождение эукариот
Следующий важнейший рубеж в эволюции жизни документируется рядом находок ископаемых остатков в отложениях, имеющих возраст 0,9—1,3 млрд лет. Среди них наиболее известна ископаемая флора, обнаруженная в углистых кремнистых сланцах формации Биттер-Спрингс (возраст 0,9 млрд лет) в Центральной Австралии. Помимо одноклеточных и нитчатых форм синезеленых водорослей (по мнению Д. Шопфа, они уже относятся к современным семействам этих растений) здесь найдены прекрасной сохранности остатки одноклеточных организмов размером 8—12 мкм, в которых удалось различить внутриклеточную структуру, похожую на ядро (рис. 27).
Обнаружены также стадии деления одного из видов этих одноклеточных организмов (Glenobotrydion aenigmati-cus), напоминающие стадии митоза — способа деления эукарио-тических (т. е. имеющих ядро) клеток. Сходные ископаемые остатки содержащих ядро клеток и стадий митоза были найдены также в Калифорнии в отложениях, имеющих возраст 1,3 млрд лет, а также на реках Мироедиха (Туруханский район) и Мая (Юго-Восточная Якутия) в отложениях возрастом 0,9—1 млрд лет.
Если интерпретация всех этих ископаемых остатков правильна[38], это означает, что около 1,6—1,3 млрд лет назад эволюция организмов прошла важнейший рубеж — был достигнут уровень организации эукариот.
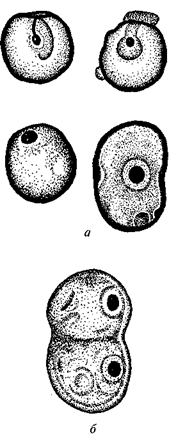
Рис. 27. Ископаемые остатки организмов из формации
Биттер-Спрингс (по Д.Шопфу):
а — Caryosphaeroides pristina; б — Glenobotrydion aenigmaticus
Ранний протерозой с полным основанием называют «веком синезеленых водорослей», получивших самое широкое распространение; это было время их расцвета. Благодаря их фотосинтетической деятельности постепенно повышалось содержание кислорода в атмосфере. Первоначально свободный кислород быстро использовался как окислитель в различных химических реакциях, в частности в процессах отложения осадочных железных руд при участии ферробактерий (основная масса железорудных толщ на всей Земле сформировалась в промежутке 2,2—1,9 млрд лет назад). После завершения этих процессов, радикально изменивших геохимический облик поверхностных слоев земной коры, кислород стал накапливаться в атмосфере во все более заметных количествах. Большие масштабы в это время приобрела и строматолитообразующая деятельность нитчатых синезеленых водорослей, которая привела к образованию огромных толщ карбонатных пород.
Возникновение эукариот — это следующий важнейший рубеж в до-кембрийской эволюции организмов после возникновения фотосинтеза. Эукариоты обладают значительно более сложной и совершенной организацией клетки, чем прокариоты. Протоплазма эукариотической клетки сложно дифференцирована: в. ней обособлены ядро и другие различные органоиды (митохондрии, пластиды, комплекс Гольджи, центриоли и др.), отделенные от цитоплазмы полупроницаемыми мембранами и выполняющие различные функции, снаружи клетка окружена клеточной мембраной. Наружная и внутренние клеточные мембраны образуют единый мембранный комплекс. Мембраны имеют сложное молекулярное строение (липидная «пленка», пронизанная белковыми «каналами»), обусловливающее избирательное проникновение веществ. У прокариот вместо клеточной мембраны клетка обволакивается единственной гигантской молекулой мукопептида (вещество, состоящее из аминокислот, углеводов и липидов). В ядре — интегрирующем центре клетки эукариот — локализован хромосомный аппарат, в котором сосредоточена основная часть кода наследственной информации. У прокариот нет такого интегрирующего центра и подобной упорядоченности наследственной информации; элементы аппарата наследственности отчасти включены в единственную так называемую «хромосому» бактериальной клетки, отчасти находятся в цитоплазме. Упорядоченность аппарата наследственности и сложность организации клетки у эукариот требуют такой же упорядоченности в передаче наследственной информации при клеточном делении. В связи с этим у эукариот развился весьма совершенный механизм клеточного деления — митоз, обеспечивающий точное и эквивалентное распределение наследственной информации в обе дочерние клетки. У прокариот митоза не наблюдается.
Одним из важнейших эволюционных достижений эукариот является возникновение механизма цитоплазматической подвижности, связанное с формированием аппарата микротрубочек (и его производных), в котором локализованы сократительные белки: миозин, актин, тубулин (у прокариот актин и тубулин встречаются редко, а миозин неизвестен). Источником энергии для функционирования сократительных белков является аденозинтрифосфат (АТФ) — основной аккумулятор и переносчик химической энергии в клетке. Возникновение митоза стало возможным именно благодаря развитию этого аппарата, с ним же связано и совершенствование механизмов активной подвижности организма во внешней среде[39].
Эукариотный уровень организации обеспечивает более эффективное выполнение всех клеточных функций и открывает перспективы для дальнейшей прогрессивной эволюции.
Имеются две основные точки зрения на проблему происхождения эукариотической клетки и соответственно эукариот как одной из высших таксономических групп. Первая из них — так называемая аутогенная концепция — предполагает постепенную дифференциацию прокариотической клетки, в ходе которой развился мембранный комплекс. Сначала могла возникнуть наружная клеточная мембрана, затем ее локальные впячивания внутрь клетки образовали внутриклеточные мембраны. На основе дифференциации мембранного комплекса структурно оформились клеточные органоиды. Согласно аутогенной концепции эукариоты возникли от одной из групп прокариот (от какой именно — указать сейчас невозможно).
Вторая точка зрения (симбиогенная концепция) получила широкую известность после работ Л.Маргулис (Саган), в 1967— 1971 гг. обосновавшей гипотезу о возникновении эукариотической клетки из симбиоза разных прокариот: крупной клетки — хозяина — и более мелких организмов, поселившихся в цитоплазме первого. Последние дали начало различным органоидам. Аргументы в пользу этой точки зрения сводятся к наличию в некоторых органоидах эукариотической клетки (митохондрии, пластиды) собственной ДНК, некоторому сходству нуклеотидных последовательностей этих органоидов с таковыми ряда прокариот, а также существованию сложных симбиотических комплексов среди современных организмов (известно, например, около тысячи видов-эндосимбионтов, обитающих в организме одноклеточных простейших животных). Согласно симбиогенной концепции предками эукариот должны быть сразу несколько групп прокариот.
Симбиогенная концепция получила широкое распространение в течение двух последних десятилетий. Однако все же аутогенная (классическая) точка зрения представляется в настоящий момент лучше обоснованной и согласующейся с общими тенденциями прогрессивной эволюции организмов (см. ч. IV). ДНК органоидов могла возникнуть путем сегрегации (обособления) фрагментов первоначально единого генома.,
Для эукариот характерен половой процесс, сущностью которого является обмен наследственной информацией между организмами, принадлежащими к одному биологическому виду. Как мы видели выше, половой процесс объединяет наследственную информацию, присущую разным индивидам, в единый видовой генофонд и, с другой стороны, обеспечивает перекомбинацию наследственной информации в каждом следующем поколении. Половой процесс известен и у некоторых прокариот (бактерии), но у них он неупорядочен и не обеспечивает эквивалентного обмена разных особей наследственной информацией.
Вероятно, практически одновременно с развитием у эукариот полового процесса у них должен был сформироваться и мейоз, необходимый для редукции числа геномов, удваивающегося при половом процессе. Мейоз также способствует возрастанию комбинативной изменчивости вида. Результатом ее резкого усиления у эукариот было значительное убыстрение их эволюции. Вместе с тем организация клеточного ядра и аппарата клеточного деления у эукариот обеспечивает гораздо более совершенную защиту генома от горизонтального переноса генов, чем это характерно для прокариот. Горизонтальный перенос генов для эукариот теряет существенное значение как фактор эволюции.
Уже около 0,9 млрд лет назад произошло разделение эволюционного ствола древнейших эукариот на ряд ветвей, давших начало разным типам водорослей: зеленым (Chlorophyta), бурым (Phaeophyta), красным (Rhodophyta) и др., а также грибам (Fungi). Д. Шопф полагает, что описанные выше ископаемые остатки различных эукариот из местонахождений возраста 0,9—1,3 млрд лет принадлежат ко всем этим группам низших растений.
Вероятно, в это время существовали уже и одноклеточные животные — простейшие (Protozoa), либо возникшие от общего предкового ствола всех эукариот, либо обособившиеся от ранних представителей одной из групп растений. От какой именно группы — с уверенностью сказать сейчас невозможно: разные исследователи связывали происхождение животных с каждой из названных групп растений. По общей совокупности данных, к предкам Protozoa наиболее близкими кажутся некоторые одноклеточные зеленые водоросли. Не случайно таких жгутиконосных одноклеточных, как эвгленовые, вольвоксовые и др., способных и к фотосинтезу и к гетеротрофному питанию, ботаники традиционно рассматривают в составе типа зеленых водорослей, а зоологи — в составе типа простейших животных.
Обращает на себя внимание отсутствие ископаемых остатков животных в отложениях большей части криптозоя. Остатки некоторых одноклеточных простейших (радиолярии и, возможно, фораминиферы) и ряда групп многоклеточных животных появляются лишь в вендских отложениях. Это может быть связано и с более поздним возникновением животных, и с худшими возможностями фоссилизации (т. е. перехода в ископаемое состояние) их клеток, лишенных в отличие от растительных прочной целлюлозной оболочки. Скелета же протерозойские животные не имели.
Дата добавления: 2015-04-16; просмотров: 268; Мы поможем в написании вашей работы!; Нарушение авторских прав |