КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
И каскадные взаимодействия
В целостном организме различные органы и структуры взаимосвязаны различными формами корреляций и координации, и цепи этих взаимосвязей сложно переплетаются друг с другом. При этом в организации многих групп организмов возникают своего рода узкие места — такие ключевые морфофункциональные особенности организации, которые через каскад различных коррелятивных и координационных взаимосвязей оказывают определяющее влияние на особенности функционирования многих зависимых систем организма.
В координационном каскаде взаимосвязанных систем состояние каждого последующего уровня определяется предшествующим, начиная от ключевой, или лимитирующей, системы. Преобразования ключевой системы могут оказывать как негативное («запрещающее»), так и позитивное («разрешающее») воздействие на эволюционные изменения зависимых систем организма. В первом случае они обусловливают возникновение морфофизиологических эволюционных запретов, препятствующих развитию определенных адаптации и направлений эволюционных преобразований, во втором, наоборот, снимают подобные запреты и ограничения, существовавшие прежде.
Очень интересны каскадные Морфофизиологические взаимосвязи в организации земноводных, определяемые особенностями их дыхательной системы. Земноводные используют для вентиляции легких нагнетательный насос, образованный подъязычным аппаратом и его мускулатурой и представляющий собой модифицированный жаберный насос предков наземных позвоночных — кистеперых рыб. Несовершенство этого дыхательного насоса, расположенного впереди легких (при отсутствии эффективных механизмов, способных изменять объем самих легочных мешков), приводит к неполноте опорожнения легких при выдохе, к сохранению там некоторого количества застойного воздуха, к смешению отработанного и свежего воздуха в ротоглоточной полости (рис. 108).
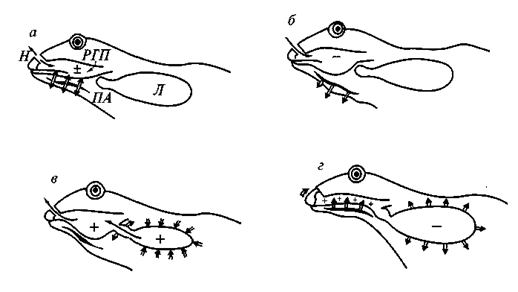
Рис. 108. Механизм вентиляции легких у бесхвостых амфибий (схема):
а — вентиляция ротоглоточной полости; б — первая фаза вдоха; в — выдох; г — вторая фаза вдоха. Значками «+» и «-» указано соотношение давления в разных отделах дыхательной системы с атмосферным давлением. Тонкие стрелки показывают направления движения воздуха в дыхательных путях, двойные стрелки — движения элементов дыхательной системы. Н— ноздри; Л — легкие; ПА — подъязычный аппарат; РГП — ротоглоточная полость
В результате хотя легкие у земноводных могут обеспечить организм достаточным количеством кислорода, но они неспособны эффективно освобождать его от углекислого газа. Это сделало необходимым развитие у земноводных дополнительного пути выведения углекислого газа — через кожу. Как мы уже упоминали, дыхательная функция кожи требует постоянной влажности по-кровов за счет деятельности кожных желез и запрещает интенсивное ороговение эпидермиса. Это резко ограничивает круг доступных для земноводных местообитаний как на суше (где амфибии активны, как правило, лишь при повышенной влажности), так и в воде: из-за высокой проницаемости покровов земноводные не могут осуществлять эффективную осморегуляцию в гипертонической среде соленых водоемов (где их организм обезвоживается, теряя влагу через покровы), и море для земноводных недоступно. Напротив, в гипотонической среде пресных водоемов через покровы в организм земноводных непрерывно поступает избыточное количество воды, которую должны удалять из организма органы выделения. Это препятствует развитию приспособлений для экономии воды в выделительной системе амфибий. Кровеносная система земноводных должна обслуживать два органа газообмена — легкие и кожу, причем кровь от них поступает к сердцу по разным магистральным сосудам (легочная вена, впадающая в левое предсердие, и кожные вены, несущие кровь через систему полых вен в правое предсердие). Это определяет необходимость смешивания обоих потоков крови в желудочке сердца и невозможность эффективного разделения артериальной и венозной крови. В результате этого для земноводных невозможно развитие эффективных систем терморегуляции и высокого уровня метаболизма, достижение гомойотермии для земноводных запрещено.
Очевидно, эволюцию дыхательной системы земноводных можно интерпретировать как пример инадаптивного варианта эволюционных преобразований, по В. О. Ковалевскому (см. с. 135). В организации земноводных несовершенный способ вентиляции легких оказался ключевым эволюционным запретом для развития важных адаптации во многих зависимых системах организма.
Но нередко и весьма совершенное само по себе приспособление может сыграть роль ключевого запрета по отношению к развитию определенных адаптации других систем организма. Эффектный пример такого рода представляют каскадные взаимосвязи в организации насекомых, также определяемые особенностями их дыхательной системы. Органы дыхания насекомых образованы сложной системой трахей — разветвленных воздухоносных трубочек, пронизывающих все тело; тончайшие трахейные стволики достигают практически всех клеток. По мнению М. С. Гилярова, именно совершенство трахейной системы позволило насекомым освоить широкий круг наземных местообитаний в условиях дефицита влаги. Вместе с тем трахейная система создала предпосылки для значительной интенсификации газообмена, вплоть до достижения у некоторых видов насекомых временной гомойотермии при полете.
С другой стороны, как было показано В. Н. Беклемишевым, трахейная система определенным образом ограничивает адаптивные и эволюционные возможности насекомых, накладывая отпечаток на всю их организацию. Крайне разветвленные трахеи делают ненужным участие кровеносной системы в газообмене, и ее периферические отделы в значительной степени редуцируются. Но при таком устройстве дыхательной и транспортной систем затруднено снабжение массивных органов, что ограничивает возможности увеличения размеров тела. Огромное большинство насекомых имеет мелкие размеры (у крупных современных видов жуков, палочников, стрекоз длина тела не превышает 13—15 см, лишь у некоторых тропических палочников она достигает 26 см). «Эволюционные эксперименты» насекомых в области крупных форм были неудачны: самые крупные известные насекомые, стреко-зы-меганевры, достигавшие около 70 см в размахе крыльев при длине тела около 30 см, вымерли еще в раннепермское время (см. с. 196).
Таким образом, у насекомых специфические особенности распределительной и дыхательной систем стали причиной возникновения эволюционного запрета на увеличение размеров тела. Небольшие размеры тела, в свою очередь, определяют многие особенности организации, поведения и экологии насекомых, которые достигли высокого совершенства в «мире малых форм». Малые размеры тела ограничивают количество клеток в организме, в частности в центральной нервной системе. Для насекомых характерна автономизация периферических отделов нервной системы (замыкание рефлекторных дуг на уровне периферических ганглиев) и преобладание наследственно закрепленных автоматизированных форм поведения. Трахейная система ограничивает также возможности экологической экспансии насекомых: при огромном разнообразии форм и адаптации насекомые совершенно не смогли освоить наиболее древнюю и обширную среду жизни — толщу морской воды. Вероятно, это связано с высокой плавучестью тела насекомых, пронизанного густой сетью воздухоносных ходов.
Дата добавления: 2015-04-16; просмотров: 290; Мы поможем в написании вашей работы!; Нарушение авторских прав |