КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Нарушения митоза
Правильное течение митоза может быть нарушено различными внешними воздействиями: высокими дозами радиации, некоторыми химическими веществами. Например, под действием рентгеновых лучей ДНК хромосом может разорваться. Хромосомы в таком случае тоже разрываются. При этом могут возникнуть хромосомы без центромерного района. Такие хромосомы лишены способности двигаться в прометафазе и анафазе. В зависимости от того, в каком месте ядра бесцентромерная хромосома находилась накануне деления,будет складываться ее дальнейшая судьба. Если хромосома была смещена к одному из полюсов клетки, то при формировании дочерних клеток она может целиком включиться в одну из них, т. е. обе сестринские хроматиды окажутся в одном ядре. Одни хромосомы начинают двигаться быстрее, другие отстают. Отставшие хромосомы могут не включиться в формирующиеся дочерние ядра. Иногда в делящейся клетке образуется не два, а три или четыре полюса, что ведет к возникновению соответственно трех или четырех дочерних клеток. При таком делении нарушается весь слаженный механизм распределения хромосом. Метафазная хромосома, состоящая из двух сестринских хроматид, может взаимодействовать одновременно только с двумя полюсами. Если полюсов больше, то каждая хромосома вынуждена "выбирать", с какими двумя полюсами из трех или четырех ей взаимодействовать. Этот выбор совершается случайно. В результате каждая дочерняя клетка получает не весь набор хромосом, а только его часть. Клетки, получившие неполный набор хромосом, как правило, оказываются нежизнеспособными и погибают. Изучение нарушений митоза, вызванных различными факторами, с одной стороны, помогает лучше понять митотические процессы, с другой - позволяет устанавливать механизмы повреждающего действия этих факторов и, следовательно, создает условия для целенаправленного поиска методов устранения таких нарушений.

25.Гаметогенез. Мейоз.
Гаметогенез или предзародышевое развитие — процесс созревания половых клеток, или гамет. Поскольку в ходе гаметогенеза специализация яйцеклетоки сперматозоидов происходит в разных направлениях, обычно выделяют оогенез (устаревшее название — овогенез в современных изданиях по эмбриологии больше не употребляется) и сперматогенез.
Гаметогенез закономерно присутствует в жизненном цикле ряда простейших, водорослей, грибов, споровых и голосеменных растений, а также многоклеточных животных. В некоторых группах гаметы вторично редуцированы (сумчатые и базидиевые грибы, цветковые растения). Наиболее подробно процессы гаметогенеза изучены у многоклеточных животных.
Мейо́з или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированных стволовых.
С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности(переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса.
Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
Профаза I — профаза первого деления очень сложная и состоит из 5 стадий:
Лептотена или лептонема — упаковка хромосом, конденсация ДНК с образованием хромосом в виде тонких нитей (хромосомы укорачиваются).
Зиготена или зигонема — происходит конъюгация — соединение гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами и их дальнейшая компактизация.
Пахитена или пахинема — (самая длительная стадия) — в некоторых местах гомологичные хромосомы плотно соединяются, образуя хиазмы. В них происходит кроссинговер — обмен участками между гомологичными хромосомами.
Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток.
Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
К концу Профазы I центриоли мигрируют к полюсам клетки, формируются нити веретена деления, разрушаются ядерная мембрана и ядрышки
Метафаза I — бивалентные хромосомы выстраиваются вдоль экватора клетки.
Анафаза I — микротрубочки сокращаются, биваленты делятся, и хромосомы расходятся к полюсам. Важно отметить, что, из-за конъюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе.
Телофаза I — хромосомы деспирализуются и появляется ядерная оболочка.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
Профаза II — происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления, перпендикулярное первому веретену.
Метафаза II — унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку.
Анафаза II — униваленты делятся и хроматиды расходятся к полюсам.
Телофаза II — хромосомы деспирализуются и появляется ядерная оболочка.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки.


26. Половые клетки человека.
Гаметы - это половые клетки: яйцеклетки (женские гаметы) и сперматозоиды (мужские гаметы), которые обеспечивают передачу наследственной информации от родителей к потомкам. Гаметы представляют собой высокодифференцированные клетки. В процессе эволюции они приобрели свойства выполнение специфических функций. Ядра как мужских, так и женских гамет содержат одинаковую наследственную информацию, необходимую для развития организма. Однако другие функции яйцеклетки и сперматозоида разные, поэтому по строению они очень различаются.
Яйцеклетки неподвижны, шаровидной или слегка удлиненной формы. Они содержат все типичные клеточные органеллы, но по строению отличаются от других клеток, так приспособлены для реализации развития целого организма. Яйцеклетки гораздо больше, чем соматические клетки. Внутриклеточная структура цитоплазмы специфическая для каждого вида животных, чем обеспечиваются видовые (а часто и индивидуальные) особенности развития. В яйцеклетках содержатся вещества, необходимые для развития зародыша. К ним относится питательный материал (желток). В некоторых видов животных накапливается в яйцеклетках столько желтка, которые можно увидеть невооруженным глазом (икринки рыб и земноводных, яйца пресмыкающихся и птиц). Из современных животных самые яйцеклетки вселедочной акулы (29 см в диаметре). У птиц яйцом считается то, что в быту называется "желтком"; диаметр яйца страуса 10,5 см, курицы - около 3,5 см.
Например, у ракообразных они должны вырасти в виде лучей или отростков, в круглых червей - форму шарообразных или овальных телец т.д. Таким образом, половые клетки существенно отличаются от соматических клеток:
1) в половых клетках гаплоидный набор хромосом, в соматических - диплоидный;
2) в половых клетках ядерно-цитоплазматическое соотношение разное: в сперматозоидах оно высокое, в яйцеклетке - низкое,
3) форма и размеры половых клеток иные, чем в соматических;
4) половые клетки отличаются низким уровнем обменных процессов;
5) для яйцеклеток характерна цитоплазматическая сегрегация закономерный перераспределение цитоплазмы после оплодотворения). Относительно большим ядром и значительным количеством цитоплазмы.


27.Особенности репродукции человека. Оплодотворение Репродукция(размножение) человека – это физиологическая функция, необходимая для сохранения человека как биологического вида. Процесс размножения у человека начинается с зачатия (оплодотворения), т.е. с момента проникновения мужской половой клетки (сперматозоида) в женскую половую клетку (яйцеклетку).
Способность к репродукции становится возможной после полового созревания. Признаком наступления полового созревания у человека являются первые поллюции у мальчиков и первые менструации у девочек. Половая зрелость наступает у лиц женского пола в возрасте 16-18 лет, мужского – 18-20 лет. Сохраняется способность к репродукции у женщин до 40-45 лет, а у мужчин до старости. Продукция гамет у представителей обоих полов различна: зрелый семенник непрерывно вырабатывает огромное количество сперматозоидов; половозрелый яичник периодически (один раз в лунный месяц) выделяет зрелую яйцеклетку, созревающую из числа овоцитов, которые закладываются на ранних этапах онтогенеза и запасы которых убывают в течение жизни женщины. Значение того, что овоциты закладываются еще до рождения состоит в том, что потомство, появляющееся к концу репродуктивного периода, развивается из овоцитов, в которых за жизнь женщины могли возникнуть генетические дефекты.
Следствием этого является то, что у пожилых матерей относительно чаще рождаются дети с врожденными дефектами. Необходимо подчеркнуть, что основную опасность представляет не сам возраст матери, а мутагенные факторы и факторы, влияющие на развитие плода. У человека, как и у других организмов, имеющих внутреннее оплодотворение, мужские половые клетки при половом акте вводятся в половые органы женщины. Во время извержения семенной жидкости (эякуляция) у человека выделяется около 100-200 млн сперматозоидов, но только один из них оплодотворяет яйцеклетку.
ОплодотворениеВстреча женских и мужских гамет происходит в верхних отделах маточных труб. Верхних отделов маточной трубы достигает около 100 сперматозоидов. Перемещение их осуществляется благодаря собственной подвижности, а также в результате мышечных сокращений стенок полового тракта и направленного движения ресничек слизистой оболочки маточных труб. Сперматозоиды сохраняют способность к оплодотворению в течение 1-2 суток, яйцеклетки – на протяжении суток после овуляции. Оплодотворение осуществляется обычно в течение первых 12 часов после овуляции. В процессе проникновения сперматозоида через барьер фоликуллярных клеток, окружающих яйцеклетку и её оболочку, большую роль играет акросомная реакция. Вслед за проникновением сперматозоида в яйцеклетку образуется оболочка оплодотворения, препятствующая проникновению других сперматозоидов. Зигота опускается по маточным трубам и на восьмые- десятые сутки зародыш внедряется в стенку матки. Если оплодотворение не наступило, яйцеклетка удаляется из организма.

29.Этапы эмбрионального развития
Период эмбрионального развития наиболее сложен у высших животных и состоит из нескольких этапов.
Период начинается с этапа дробления зиготы, т. е. серии последовательных митотических делений оплодотворенной яйцеклетки. Образующиеся в результате деления две клетки (и все последующие их поколения) на этом этапе называются бластомерами. Одно деление следует за другим, причем не происходит роста образующихся бластомеров и с каждым делением клетки становятся все более мелкими. Такая особенность клеточных делений и определила появление образного термина «дробление зиготы»
В результате дробления (когда количество бластомеров достигнет значительного числа) образуется бластула. Часто она представляет собой полый шар (например, у ланцетника), стенка которого образована одним слоем клеток — бластодермой. Полость бластулы — бластоцель, или первичная полость, заполнена жидкостью.
На следующем этапе осуществляется процесс гаструляции — формирование гаструлы. У многих животных она образуется путем впячивания бластодермы внутрь на одном из полюсов бластулы при интенсивном размножении клеток в этой зоне. В результате и возникает гаструла.
Наружный слой клеток получил название эктодермы, а внутренний — энтодермы. Внутренняя полость, ограниченная энтодермой, полость первичной кишки сообщается с внешней средой первичным ртом, или бластопором. Существуют и другие типы гаструляции, но у всех животных (кроме губок и кишечнополостных) этот процесс завершается образованием еще одного клеточного пласта — мезодермы. Она закладывается между энто- и эктодермой.
По завершении этапа гаструляции появляются три клеточных пласта (экто-, эндо- и мезодерма), или три зародышевых листка.
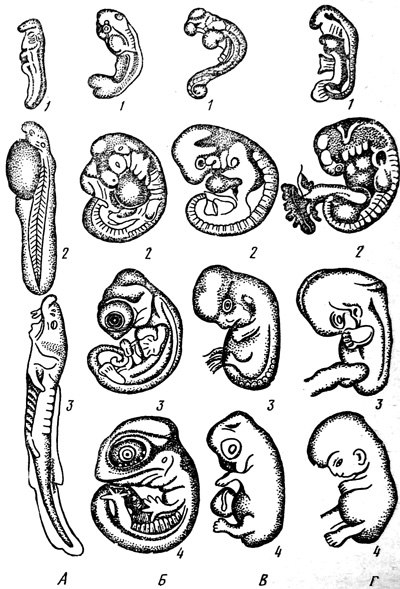
30.Критические периоды эмбриогенеза человека. Тератогенные факторы.
Критические периоды эмбриогенеза это временные промежутки, характеризующиеся повышенной чувствительностью плода к влиянию повреждающих факторов.
Критические, потому что вероятность возникновения какой-либо патологии в эти периоды очень высока.
В эмбриональном развитии человека имеются следующие3 основных критических периода.
1. Имплантация (6 – 7-е сутки после зачатия) – внедрение зиготы в стенку матки.
2. Плацентация (конец 2-й недели беременности) – образование у эмбриона плаценты.
3. Перинатальный период (роды) – переход плода из водной в воздушную среду через 9 месяцев после зачатия.
С критическими периодами в организме новорожденного связаны резкое изменение условий существования и перестройка деятельности всех систем организма (изменяется характер кровообращения, газообмена, питания). Наиболее уязвимым эмбрион становится в первом и втором критических периодах онтогенеза.
Существует масса неблагоприятных факторов, вызывающих вышеперечисленные нарушения у плода. Это, например, интоксикация материнского организма, диабетическая и алкогольная фетопатии, заболевания эндокринной системы, воздействие физических (температура, ионизирующее излучение, механическое воздействие), химических (медикаменты, алкоголь, курение, наркотики, пары ртути, толуол, хлордифенил) и биологических (инфекционные болезни, вирусы) факторов. Такие факторы называются тератогенными - факторы, которые могут привести к нарушению развития плода и его органов.
Основные группы тератогенных факторов.
1. Лекарственные средства и химические вещества.
2. Ионизирующее излучение.
3. Инфекции.
4. Метаболические нарушения и вредные привычки у беременной.


31.Старение как этап онтогенеза. Теории старения.
Старение - это универсальный и закономерный процесс, которому присуща последовательность, прогрессирование, которое приводит к снижению адаптационных возможностей и жизнеспособности индивида. Старение касается всех уровней организации: от молекулярных структур до целостного организма.
К наиболее характерным:
1. Уменьшение роста (на 0,5-1 см за пятилетие после 60)
2. Изменение формы тела (сглаживание контуров, усиление кифоза, перераспределение жирового компонента)
3. Снижение массы тела, упругости и эластичности мышц
4. Снижение амплитуды движений грудной клетки
5. Снижение плавности движений
6. Уменьшение размеров лица впоследствии потери зубов и редукции альвеолярных отростков челюстей
7. Увеличение объёма мозговой части черепа, ширины носа и рта
8. Изменения в коже (уменьшение количества сальных желез, толщины эпидермиса)
Для процесса старения характерны также изменения в функционировании важных систем организма:
1. В ЦНС наблюдаются структурные (уменьшение массы мозга, величины и плотности нейронов) и функциональные (снижение работоспособности нейронов)
2. Снижение остроты зрения, функции слухового аппарата, вкуса, части кожной чувствительности)
3. Уменьшение массы желез, снижение их гормонообразовательной функции (щитовидная, половых желез)
4. Снижается секреторная активность пищеварительных органов, жизненная ёмкость лёгких, основных функций почек, сокращающая целостность миокарда, ритмическая деятельность сердца уменьшается
5. Резко снижается иммунный гомеостаз, количество и функциональная активность Т-лимфоцитов (снижение активности иммунной системы приводит к развитию аутоиммунных процессов, растёт возможность образования опухолей)
Старение - длительный процесс, последним этапом которого является старость.
Французский биолог Ж.Бюффон рассчитывал, что длительность жизни человека должна превышать длину его роста в 6-7 раз и составляет около 90-100 лет. Все последующие расчёты по разным критериям были близки к этой цифре(максимальная длительность жизни человека достигает 130 лет).
У человека различают:
- физиологическую старость, связанную с календарным возрастом
- преждевременноесатрение, обусловленное социальными факторами и болезнями.
60-75 лет - пожилой
74 лет и более - старость
Теории старения:
1) одна из наиболее известных первых теорий: теория немецкого врача Х.Гуфеланде, который придавал в долголетии значение трудовой деятельности. Он говорил, что ни один лентяй не дожил до преклонного возраста.
2) эндокринная теория Бертольда, который показал, что пересадка семенников от одних животных к другим сопровождается развитием вторичных половых признаков. В начале 20в. уже сложилось убеждение о том, что наступление старости связано с угасанием деятельности желез внутренней секреции, в частности половых желез. На основе этого убеждения в разных странах было проделано много операций по омолаживанию пожилых или старых людей. Все эти операции приводили к некоторым эффектам, но лишь временным. После таких воздействий процессы старения продолжались, причём ещё интенсивнее.
3) микробиологическая теория старения И.И.Мечникова, различавшего физиологическую старость и патологическую. Он считал, сто старость человека является патологической, т.е. преждевременной. Основу представлений И.И.Мечникова составляло учение об ортобиозе, в соответствии с которой основной причиной старения является повреждение нервных клеток продуктами интоксикации, образующимися в результате гниения в толстьм кишечнике. Развивая учение о нормальном образе жизни (соблюдение правил гигиены, регулярный труд, воздержание от вредных привычек), И.И.Мечников предлагал также способ подавления гнилостных бактерий кишечника путём употребления кисломолочных продуктов.
4) теория о роли ЦНС в старении И.П.Павлова, который утановил интегрирующую роль ЦНС в нормальном функционировании организмов. Последователи И.П.Павлова в экспериментах на животных показали, что преждевременное старение вызывается нервными потрясениями и продолжитьным нервным потрясением.
5) теория возрастных изменений соединительной ткани, сформулированная А.А.Богомольцем.он считал, что физиологическую активность организма обеспечивает соединительная ткань (костная, хрящи, сухожилия, связки и волокнистая соединительная ткань) и что изменения строения клеток определяют возрастные изменения.
Наиболее распространенные современные представления о механизме старения сводятся к тому, что в процессе жизнедеятельности в клетках организма накапливаются соматические мутации, в результате которых происходит синтез дефектных белков или нерепарируемые сшивки ДНК с белком. Поскольку дефектные белки играют деезинтегрирующую роль в клеточном метаболизме, то это ведёт к старению. Однако исчерпывающей теории старения все же не создано, поскольку ясно, что ни одна из этих теорий самостоятельно объяснить механизмы старения не может.

33.Клиническая и биологическая смерть.
Клиническая смерть – это период, когда все угасшие жизненно важные процессы организма еще обратимы. Клиническая смерть может быть вызвана обильным кровотечением, электротравмой, утоплением, рефлекторной остановкой сердца, острым отравлением и т.д.
Признаки клинической смерти:
· отсутствие пульса на сонной или бедренной артерии(основной признак остановки кровообращения),
· отсутствие дыхания,
· потеря сознания,
· широкие зрачки и отсутствие их реакции на свет.
Биологи́ческая смерть (или истинная смерть) - необратимое прекращение физиологических процессов в клетках и тканях. Связана с прекращением самообновления, химические процессы становятся неупорядоченными, в клетках происходит аутолиз (самопереваривание), разложение. Скорость процессов определяется степенью чувствительности. Наиболее чувствительные - нервные клетки (изменения происходят уже через 5-6 минут).
К ранним признакам биологической смерти относятся:
· Отсутствие реакции глаза на раздражение (надавливание)
· Помутнение роговицы
· Появление симптома «кошачьего глаза»: при боковом сдавлении глазного яблока зрачок трансформируется в вертикальную веретенообразную щель, похожую на кошачий зрачок.
· Отсутствие пульса.
В дальнейшем обнаруживаются трупные пятна с локализацией в отлогих местах тела, затем возникает трупное окоченение, разложение. Трупное окоченение обычно начинаются с мышц лица, верхних конечностей.


34.Регенерация и её виды.
Регенерация – процесс восстановления утраченных или поврежденных тканей или органов.
Различают физиологическую, репаративную и патологическую регенерацию.
Физиологическая регенерация свойственна всем организмам. Процесс жизнедеятельности обязательно включает два момента – утрату и восстановление морфологических структур на клеточном, тканевом, органном уровнях. У млекопитающих и человека непрерывно слущиваются наружные слои кожного эпителия, продолжительность жизни кишечного эпителия составляет несколько дней. Сравнительно быстро происходит смена эритроцитов.
Репаративная регенерация возникает, когда в организме происходит повреждение и гибель клеток и тканей. Она широко распространена, но способность к ней выражена неодинаково у различных животных. Репаративная регенерация может быть типичной(гомоморфоз) и атипичной(гетероморфоз). При гомоморфозе восстанавливается такой же орган, как и утраченный, при гетероморфозе восстановленные органы отличаются от типичных.
Восстановление осуществляется путем эпиморфоза, морфаллаксиса и эндоморфоза.
Эпиморфоз – рубцевание, безвосстановление органа. Морфаллаксис – разрастание оставшейся части, завершается образованием организма или органа. Эндоморфоз (регенерационная гипертрофия) – восстановление внутри органа.
Патологическая регенерация – разрастание тканей, не идентичных здоровым тканям в данном органе. Например, на месте глубоких ожогов может быть массивное разрастание плотной соединительной рубцовой ткани, а нормальная структура кожи не восстанавливается.

40.Взаимодействие аллельных и неаллельных генов
Взаимодействиеаллельныхгенов:
1) Полное доминирование - взаимодействие аллельных генов, при котором доминантный ген полностью подавляет фенотипическое проявление рецессивного гена. Например, жёлтый цвет гороха полностью подавляет зелёный.
2) Неполное доминирование - когда оба аллельных гена и доминантный, и рецессивный проявляются фенотипически в результате чего гибридное потомство имеет промежуточный фенотип. Например, у цветов АА - красный, аа - белый, а Аа - розовый.
3) Кодоминантность - вид взаимодействия, когда оба гена проявляют каждый себя самостоятельно в равной степени.
4) Сверхъдоминирование - заключается в том, что у доминантного аллеля в гетерозиготном состоянии иногда отмечается более сильное проявление, чем в гомозиготном состоянии.
Взаимодействие неаллельных генов:
1) Комплементарное действие - когда для формирования признака необходимо наличие двух или более неаллельных (обычно доминантных) генов. Например, у человека нормальный слух обусловлен двумя доминантными генами Д и Е, из которых один определяет развитие улитки, а другой - слухового нерва. Доминантные гомозиготы и гетерозиготы по обоим генам имеют нормальный слух, рецессивные по одному из этих генов - глухие.
2) Эпистаз - подавление неаллельным геном действия другого гена, названного гипостатическим.
3) Полимерия - заключается в том, что различные неаллельные гены могут оказывать действие на один и тот же признак, усиливая его проявление. Такие гены получили название полимерных, а признаки ими определяемые - пол генных. Так цвет кожи у человека определяется 5ю парами аллельных генов.
4) Плейотропия - зависимость нескольких признаком от одного гена, т.е. проявление множественных эффектов одного гена, явление обратное полимерии. Например, болезнь Марфана, арахнодактилия - наследственноезаболение человека. Гены, определяющий это заболевание, вызывают нарушение развития соединительной ткани и оказывает влияние одновременно на развитие нескольких признаков: нарушение в строении хрусталика глаза, аномалии в ССС.

\ 
41.Сцепленное наследование
В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила.
Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде.

Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.

1 — некроссоверные гаметы; 2 — кроссоверные гаметы.
Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом.
Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом.
Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.

| Образуются гаметы: | 
|
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правилокроссоверные гаметы составляют небольшую часть от всего количества гамет.

| Образуются гаметы: | 
|
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
43.Генетика пола
В хромосомном наборе у женщин имеется 22 пары аутосом и две одинаковые половые хромосомы XX; у мужчин 22 пары аутосом и половые хромосомы X и V (неодинаковые). В процессе мейоза каждая из пары гомологичных хромосом уходит в разные гаметы. Так как у женщин 23 пары гомологичных хромосом, то во все гаметы попадает 22 аутосомы и одна Х-хромосома (гаметы одинаковы), поэтому женский пол гомогаметный. У мужчин образуется два типа гамет: 22+Х и 22+Y, поэтому мужской пол гетерогаметный. Вероятность рождения девочек так же, как и мальчиков, составляет 50 %.
P - ♀22×2 + XX × 22×2 + XY♂
G – 22+X 22+X 22+Y
F - 22×2 + XX 22×2 + XY
50% девочки 50% мальчики
Пол будущего ребенка определяется сочетанием половых хромосом в момент оплодотворения. Если яйцеклетку оплодотворяет сперматозоид с Х- хромосомой, то рождается девочка, а если яйцеклетку оплодотворяет сперматозоид с Y-хромосомой, то рождается мальчик. Признаки организма, связанные с полом, подразделяют на три категории.
1)признаки, сцепленные с половыми хромосомами. Это признаки, развитие которых обусловлено генами, расположенными в одной из половых хромосом. Х - хромосома по размерам значительно больше, чем Y - хромосома.
В Х-хромосоме есть большой участок, которого нет в Y-хромосоме, в нем расположены рецессивные гены (гемофилии, дальтонизма, миопатии и доминантные (рахита, не поддающиеся лечению витамином D, темной эмали зубов). Гены этого участка полностью сцеплены с полом, передаются исключительно через Х-хромосому. Доминантные гены из этого участка одинаково проявляются у обоих полов; рецессивные — у женщин только в гомозиготном состоянии, а у мужчин - в гемизиготном состоянии.
Рассмотрим наследование, сцепленное с полом, на примере гемофилии. У мужчин ген гемофилии находится в гемизиготном состоянии (один из пары), и поэтому гемофилия у мужчин всегда проявляется. У женщин, если и есть, этот ген в гетерозиготном состоянии, он не проявляется, так как доминантный ген нормальной свертываемости его подавляет. В брак вступают здоровая женщина-носительница и здоровый мужчина:
H-ген нормальной свертываемости крови
h-ген гемофилии
P-♀ XHXh × XHY♂
G- XHXhXHY
F - XHXH ,XHXh , XHY, XhY
XHXH -девочка здоровая
XHXh - девочка носитель
XHY - мальчик здоровый
XhY - мальчик больной
В непарном участке У-хромосомы, кроме генов, детерминирующих пол, расположено небольшое число генов, которые могут встречаться только у лиц мужского пола и передаются от отца всем сыновьям (голандрические гены): волосатость ушей, перепонки между пальцами ног, ихтиоз (кожа в виде рыбьей чешуи).
В парном сегменте (гомологичном для Х- и У-хромосом) локализованы гены, детерминирующие пигментную ксеродерму, болезнь Огучи, спастическую параплегию, эпидермолиз буллезный, общую цветовую слепоту и другие. Их "называют неполно или частично сцепленными с полом. Они могут передаваться как с Х-, так и с У-хромосомой;
2) признаки, контролируемые полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но степень проявления их (экспрессивность) разная у мужчин и женщин (особенно у гетерозигот, так как происходит сдвиг доминантности). Изменение доминирования гена обусловлено половыми гормонами. Например: а) у мужчин ген облысения доминантный, а у женщин — рецессивный и проявляется только в гомозиготном состоянии, поэтому лысых мужчин значительно больше, чем женщин; б) пенетрантность гена подагры у мужчин 80 %, а у женщин 12 %, значит, подагрой чаще болеют мужчины;
3) признаки, ограниченные полом. Развитие этих признаков обусловлено генами, расположенными в аутосомах обоих полов, но проявляются они только у одного пола (гены удойности и жирности молока имеются у коров и быков, но проявляются только у коров).



44.Изменчивость и ее формы
Изменчивость – способность организмов приобретать новые или утрачивать прежние признаки или свойства.
Степень фенотипического проявления данного гена называется экспрессивностью, а частота его проявления - пенетрантностью. С изменчивостью связаны явления фенокопий и генокопий. Фенокопии — изменения фенотипа под влиянием неблагоприятных факторов среды, по проявлению похожие на мутации. В медицине фенокопии — ненаследственные болезни, сходные с наследственными.Генокопии—это одинаковые изменения фенотипа, обусловленные аллелями разных генов.
Изменчивость
1. Ненаследственная, модификационная или фенотипическая;
2. Наследственная, мутационная или генотипическая;
3. Наследственная, комбинативная или генотипическая.
Модификационная изменчивость возникает под действием окружающей среды. По наследству не передаётся, потому что при модификациях меняется только фенотип (признак), а генотип не меняется.
Например, если человек будет находится на солнце, то он загорит;
Границы, внутри которых могут происходить модификационные изменения, называются «норма реакции», они заложены в генотипе.
Дата добавления: 2015-01-19; просмотров: 774; Мы поможем в написании вашей работы!; Нарушение авторских прав |