КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Отношения хищник-жертва, паразит-хозяин. Математические модели. Эксперименты А. Ф. Гаузе.
Хищник – жертва.
В основе этой формы биоценотических отношений лежат прямые трофические связи. Большую роль в пищедобывательной деятельности животных-зоофагов играет поведение. Их пища – подвижные формы со своими особенностями биологии и поведения. Поэтому типы охоты у разных плотоядных животных хорошо коррелируют со спецификой биологии жертв. Так, если жертвы многочисленны, не прячутся в убежища и относительно малоподвижны, характер охоты имеет вид простого собирательства (питание планктонными животными, образующими в толще воды большие скопления). При таком типе питания нередки миграции потребителей вслед за передвижениями больших скоплений планктона. Если добыча достаточно многочисленна, но ведёт скрытый образ жизни, наблюдается охота типа «широкого поиска», когда животное всё время перемещается, обшаривая по пути укрытия, в которых может находиться пища. Так ведут себя кормящиеся стайки синиц и некоторые других насекомоядных птиц. При охоте на относительно крупную и немногочисленную добычу характерно подстерегание или высматривание жертвы с места, дающего хороший обзор. В обоих случаях охота состоит в преследовании конкретной особи - жертвы. Так добывают пищу осьминоги, некоторые хищные рыбы, соколы, гепарды и т.д.
Приспособления жертв могут быть морфологическими (твёрдые покровы, шипы, колючки и пр.), поведенческими (затаивание, убегание, активная оборона) или физиологическими (продукция ядовитых или отпугивающих веществ). Эксперименты в природе показали, что вторичные метаболиты кораллов р. Sinularia оказывают эффективное отпугивающее действие на рыб. Аналогичный способ защиты известен для многих насекомых и их личинок; у ряда видов он сочетается с предупреждающим типом окраски. На этой основе может возникать особая форма мимикрии, при которой у нескольких несъедобных видов окраска оказывается сходной – так называемая «мюллеровская» мимикрия. Биологический смысл её в том, что в этом случае выработка у хищника негативной реакции происходит при меньшем уровне гибели для каждого из сходно окрашенных видов. Другой тип мимикрии – подражание несъедобным объектам со стороны видов, не обладающих защитными механизмами. Пример: имитация бабочкой Liminitis archippus (вице-король) несъедобного вида – бабочки монарх Danaus plexipus.
Хищники тоже используют ядовитые вещества, в частности для умерщвления своих жертв. Например, ядовитые змеи (гремучая змея Crotalus engo). Реже ядовитые вещества используются для обездвиживания жертв. В составе секрета слюнных желёз короткохвостой землеройки Blarina brevicauda содержится яд замедленного действия, парализующий насекомых, которые после этого ещё 3 – 5 дней остаются живыми. Благодаря этому землеройка может иметь запас «живых консервов».
Некоторые животные способны менять окраску в зависимости от цвета фона. Например, головоногие моллюски, некоторые ракообразные, рыбы, амфибии, рептилии. Экспериментально показано, что из всех рыб, выживших в водоёме, где охотился пингвин, 66% составили особи, окрашенные соответственно фону.
Взаимодействия хищников и их жертв очень динамичны и взаимообусловлены. Это особенно ярко выражается в проявлениях адаптивного поведения. Наблюдения за головастиками американской жабы Bufo americans показали, что в присутствии хищных личинок стрекоз они снижают активность, изменяется и их распределение в пространстве. Например, у коловраток Brahionus calchforus в присутствии хищной коловратки Asplanchna развиваются длинные боковые выросты панциря, которые препятствуют заглатыванию их. Образование этих шпор индуцируется веществом выделяемым хищником и уменьшением пищи.
Результатом воздействия хищников на популяции жертв нередко является изменение их демографической структуры. Это связано с выборочностью питания. «Санитарная» роль хищников – добывание больных, слабых и старых животных. Избирательность в охоте приводит и к сдвигам полового и возрастного состава популяции жертв. Например, в добыче волков старые (>=9,5 лет) лоси составили 57%, тогда как среди отстреленными охотниками – всего 7 %.
Параллельное развитие противоположно направленных адаптаций в системе «хищник – жертва» ведёт к тому, что ни одно из приспособлений не становится абсолютным, но их взаимодействие определяет поддержание такого уровня изъятия хищниками их жертв, при котором возможно длительное сосуществование популяций тех и других в составе экосистем.
Паразит – хозяин.
Паразитизм – это форма взаимоотношений двух различных видов организмов, носящих онтогенетический характер, при которой организмы одного вида (паразита, потребителя) живут за счёт питательных веществ или тканей организма другого вида (хозяина) в течение определённого времени. Паразитизм известен на всех уровнях организации живого, начиная с вирусов и бактерий и заканчивая высокоорганизованными животными.
Распространение паразитизма. Чем сложнее устроен организм, тем выше эффективность его гомеостазирующих систем и поэтому больше благоприятных возможностей он предоставляет в качестве среды обитания. Чем организм совершеннее, тем меньше становится для него потребность использовать благоприятные условия в другом организме. Это объясняет, почему явление паразитизма наиболее представлено среди микроорганизмов и примитивных многоклеточных. Число видов, ведущих паразитический образ жизни, резко падает у высокоорганизованных организмов. У высших животных встречается наиболее обильное разнообразие населения паразитов.
Формы паразитизма и взаимные адаптации паразитов и их хозяев.
· Эктопаразитизм (клещи, напр. таёжный клещ, собачий клещ; блохи);
· Эндопаразитизм (паразитические черви, напр. печёночный сосальщик, широкий лентец).
По степени тесноты связей паразита и хозяина выделяют 2 формы:
1. Облигатный – ведёт только паразитический образ жизни, не выживает без обязательной связи с хозяином (все черви паразиты, напр. Taeniarhynchus saginatus – бычий цепень). Характеризуется наиболее полным комплексом приспособлений к использованию организма хозяина как среды.
2. Факультативный – ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому существованию. Например, Armillaria mellea – опёнок. Их связь исключительно трофическая. Морфофизиологические и биологические адаптации к паразитизму выражены слабо.
Адаптации парзитов: среда в виде живого и подвижного организма требует формирования способов фиксации в ней: зацепки, крючья, присоски, видоизменения формы тела. Эктопаразиты имеют сплющенное тело. У эндопаразитов утрачиваются органы, которые не нужны им в среде хозяина.
Математические модели.
При увеличении плотности популяции резко возрастает на неё влияние хищников и паразитов. Взаимоотношение «хищник – жертва», «паразит – хозяин», или «потребитель – ресурс» наиболее полно показывают принцип действия внутренних факторов. Так, в однородной (гомогенной) среде, не имеющей укрытий для размножения, хищник рано или поздно уничтожает популяцию жертвы и после этого вымирает сам.
В естественных условиях (в гетерогенной среде) возникает следующая временная и причинно-следственная цепь: размножение жертвы→размножение хищника→резкое сокращение численности жертвы→падение численности хищника→размножение жертвы и т.д.
Волны жизни хищника и жертвы следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника так и жертвы остаётся примерно на одном уровне. Длительность периода зависит от скорости роста обоих видов, а так же от исходных параметров. Для популяции жертвы влияние хищника положительно, т.к. её чрезмерное размножение привело бы к краху её численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.
 ГРАФИКИ. Взаимоотношения между хищником и жертвой: А – в гомогенной среде инфузория Dinidium nasatum погибает от голода после поедания жертвы; Б – в гетерогенной среде – жертва частично может укрыться от хищника, и он вымирает; В – естественные колебания численности хищника и жертвы ( по Гаузе).
ГРАФИКИ. Взаимоотношения между хищником и жертвой: А – в гомогенной среде инфузория Dinidium nasatum погибает от голода после поедания жертвы; Б – в гетерогенной среде – жертва частично может укрыться от хищника, и он вымирает; В – естественные колебания численности хищника и жертвы ( по Гаузе).
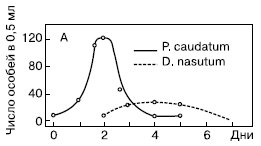
Рис. 3. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в однородной среде (количество особей в 0,5 см3)

Рис. 5. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде (количество особей в 0,5 см)

Рис. 6. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде при наличии иммиграций (1 инфузория туфелька + 1 дидиниум)
Эксперименты Гаузе.
Жертва – инфузория туфелька Paramecium caudatum.
Хищник – инфузория дидиниум Didinium nasutum.
В качестве пищи для инфузории туфельки Гаузе использовал культуру сенной палочки (Bacillus subtilis).
Первая серия опытов была поставлена Гаузе в так называемой однородной среде. Для этого использовалась разведенная бактериальная культура без осадка. Особенность биологии дидиниум заключается в том, что он не является активным охотником, а просто хватает все, что попадается ему на пути. Инфузория туфелька, ускользая от хищника, использует значительно бóльшую скорость передвижения, а также особые защитные образования – трихоцисты. При этом она никогда не прячется в убежища. Совокупность этих особенностей определила чистоту данной серии опытов и позволило Гаузе выделил процесс взаимодействия хищника и жертвы в чистом виде. Бактериальная культура без осадка разливалась в пробирки объемом 0,5 мл. В нее помещалось 5 особей инфузории туфельки, а через 2 суток вносили 3 особи хищной инфузории дидиниум. Практически сразу же после внесения хищников в культуру, численность жертвы начинала убывать, поскольку дидиниум поедает новую инфузорию туфельку через каждые 3 часа. Дидиниум интенсивно размножался, достигая максимума численности к 3-м – 4-м суткам опыта. На 5-е сутки дидиниум выел всех инфузорий туфелек, а на 6-е – все хищники погибли от голода.
Из проведенной серии опытов Гаузе сделал вывод, что в однородной среде при постоянных внешних условиях в системе «хищник–жертва» отсутствуют собственные периодические колебания численности видов.
Во второй серии опытов Гаузе усложнил условия эксперимента. Желая немного приблизиться к природным условиям, он создал в среде «убежище», где могли бы скрываться инфузории туфельки. Для этого Гаузе использовал густую бактериальную среду с осадком, который и стал «убежищем». Концентрация бактерий в осадке значительно выше, чем вне его. Это привлекает инфузорий туфелек, которые постепенно концентрируются в осадке вне зависимости от того, охотится за ними дидиниум или нет. При этом, попадая в осадок, инфузории туфельки становятся недоступными для хищника. Таким образом, он представлял своеобразную полупроницаемую перегородку, через которую могли проникать инфузории туфельки, а дидиниум – нет. Важным условием этой серии опытов являлось отсутствие резких встряхиваний и перемешивания среды. В противном случае «убежище» могло быть нарушено. Густая бактериальная среда с осадком разливалась в пробирки объемом 0,5 мл. В нее одновременно вносилось 5 особей инфузории туфельки и 3 особи инфузории дидиниум. К началу 2-х суток число хищников несколько возрастало за счет выедания части жертв. Однако большая часть инфузорий туфелек, находясь в осадке, оставалось недоступной для хищника. В результате из-за недоступности жертвы (т. е. недостатка пищи) по окончании 2-х суток численность дидиниум начинала снижаться. Одновременно с этим наблюдалось интенсивное размножение инфузории туфельки. Примерно к середине 5-х суток опыта дидиниум полностью вымирал, а численность инфузории туфельки резко возрастала.
Таким образом, в результате появления в среде «убежища», некоторое количество жертв ускользает от хищника, что позволяет им в конце концов полностью занять среду. В природе наличие убежищ играет важную роль в процессе борьбы за существование.
В третьей серии опытов Гаузе использовал разведенную бактериальную среду без осадка (т. е. лишенную «убежища»). По 2 мл среды наливалось в небольшие стеклянные чашки с плоским дном. Наблюдения проводились над одной и той же культурой, без вмешательства в состав ее содержимого. В начале опыта, а затем на каждый 3-й день в культуру вносились 1 инфузория туфелька и 1 дидиниум. При этом из культуры хищного вида отбирались сильно измельчавшие особи. Если в течение следующих 12 часов они не находили жертву, то погибали.
При первой иммиграции в свободную среду дидиниум не нашел жертвы и погиб. После этого начался интенсивный рост инфузории туфельки. К моменту второй иммиграции концентрация инфузории туфельки в среде была уже очень велика, благодаря чему начался рост популяции дидиниум. Третья иммиграция пришлась на момент интенсивного уничтожения жертв хищником, и потому не внесла существенных изменений в общую картину. К четвертой иммиграции дидиниум выел всех инфузорий туфелек, после чего погиб сам. Внесенная в среду инфузория туфелька дала начало новому циклу роста жертв. В дальнейшем, такие изменения численности обоих видов регулярно повторялись.
Проанализировав результаты, полученные в ходе трех серий опытов, Гаузе пришел к заключению, что периодические колебания численности хищника и жертвы не являются свойством процесса взаимодействия между ними, как это предполагали математики. Вероятно, они возникают в результате постоянного вмешательства извне (убежища для жертв, сохранение хищников путем инцистирования, иммиграция и эмиграция, колебания факторов среды и т. д.). Именно поэтому в естественных условиях природа периодических колебаний численности находящихся в непосредственном взаимодействии видов может быть весьма сложной.
Дата добавления: 2015-04-18; просмотров: 758; Мы поможем в написании вашей работы!; Нарушение авторских прав |