КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Потенциал действия. Развитие ПД. Характеристика. Стадии.
ПД – называется электрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения.
Определяется разной концентрацией ионов по разные стороны от мембраны и диффузия ч/мембрану.
В опытах по использованию ПД использовала 2а микроэлектрода, введенных в аксон. На первый микроэлектрод подается импульс с амплитудой V от генератора Г прямоугольных импульсов, меняющих мембранный потенциал. Мембранный потенциал измеряется при помощи второго микроэлектрода высокоомным регистратором напряжения Р.
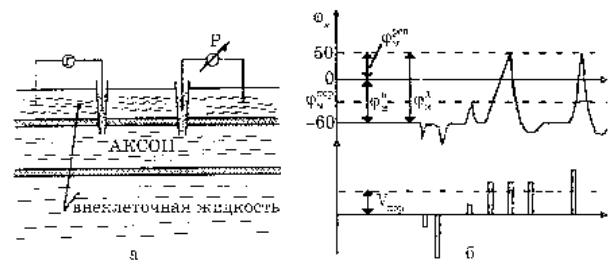 Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. В этом случае, кода возбуждающий импульс смещается еще дальше в отрицательную сторону, он сопровождается гиперполяризацией мембраны. Так же не формируется потенциал действия, гогда потенциал действия положительный (деполяризующий), но его амплитуда меньше порогового значения Vпор. Однако если амплитуда положительного, деполяризующего импульса окажется больше значения V пор, jм становится больше jмпор и в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранныйпотенциал даже меняет свой знак – становится положительным (jвн >jнар).
Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. В этом случае, кода возбуждающий импульс смещается еще дальше в отрицательную сторону, он сопровождается гиперполяризацией мембраны. Так же не формируется потенциал действия, гогда потенциал действия положительный (деполяризующий), но его амплитуда меньше порогового значения Vпор. Однако если амплитуда положительного, деполяризующего импульса окажется больше значения V пор, jм становится больше jмпор и в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранныйпотенциал даже меняет свой знак – становится положительным (jвн >jнар).
Достигнув некоторого положительного значения jм рев – потенциала реверсии, мембранный потенциал возвращается к значению потенциала покоя, совершив нечто вроде затухающего колебания. После снятия возбуждения еще в течении 1-3мс в мембране наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна.
Новый деполяризующий потенциал V>Vпор может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причем амплитуда потенциала действия не зависит от амплитуды деполяризующего потенциала. Если в покое мембрана поляризована, то при возбуждении происходит деполяризация мембраны и после снятия возбуждения происходит реполяризация мембраны.
Характерные свойства ПД:
1)наличие порогового значения деполяризующего потенциала
2) Закон «все или нечего», т.е. если деполяризующий потенциал больше порогового, развивается ПД, амплитуда которого не зависит от амплитуды возбуждающего импульса и нет ПД, если амплитуда деполяризующего потенциала меньше пороговой
3) Есть период рефактерности, невозбудимости мембраны во время развития потенциала действия и остаточных явлений после снятия возбуждения.
4) В момнт возбуждения резко уменьшается сопротивление мембраны.
38. Потенциал действия. Роль ионов Na+ и K+ в генерации потенциала действия в нервных и мышечных волокнах. Кинетика изменения потоков ионов при возбуждении.
В мембране нейрона имеются потенциал-зависимые натриевые каналы, которые открываются при изменении заряда мембраны.
При деполяризации мембраны происходит открытие натриевых каналов и увеличение натриевой проводимости. Вследствие этого увеличивается вход натрия внутрь клетки, который в свою очередь увеличивает деполяризацию мембраны. Это процесс с положительной обратной связью.Если открыто достаточное количество натриевых каналов для запуска процесса, то он дальше развивается самостоятельно и идёт до конца, генерируя потенциал действия.
Процесс реполяризации свзяан с открытием потенциал-зависимых калиевых каналов. Реполяризация обеспечивается также инактивацией натриевых каналов.
Потенциал действия подчиняется принципу «всё или ничего»: если не достигнут критический уровень деполяризации мембраны, то возникает локальный потенциал, если достигнут – возникает потенциал действия, а усиление раздражения не влияеть на его величину. Амплитуда потенциала действия не зависит от силы раздражения.
39. Биопотенциалы действия. Калиевый, натриевый и суммарный ионные токи в процессе возбуждения мембраны (графики). Блокаторы ионных потоков через мембрану.
40. Графики изменения натриевого и калиевого тока через мембрану аксона при различных значениях фиксированного трансмембранного потенциала.
41. Структура натриевых каналов, их свойства: размер, дискретность и взаимонезависимость действия, время открытого состояния, время жизни, зависимость активации от мембранного потенциала.
42. Биопотенциал действия. Уравнение Ходжкина - Хаксли. Метод фиксации потенциала.
Уравнение Ходжкина-Хаксли для ПД (токи через мембрану при её возбуждении).
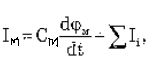 где Im – ток черех мембрану, См – емкость мембраны, å Ii – сумма ионных токов через мембрану. Электрический ток через мембрану складывается из ионных токов: ионов натрия и ионов калия и других ионов, в том числе ионов хлора, так называемого тока утечки Iут, а так же емкостного тока.
где Im – ток черех мембрану, См – емкость мембраны, å Ii – сумма ионных токов через мембрану. Электрический ток через мембрану складывается из ионных токов: ионов натрия и ионов калия и других ионов, в том числе ионов хлора, так называемого тока утечки Iут, а так же емкостного тока.
Емкостной ток обусловлен перезарядкой конденсатора, который представляет собой мембрана, перетеканием зарядов с одной ее поверхности на другую. Его величина определяется количеством заряда, перетекающего с обной обкладки на другую за еденицу времени. Полный мембраный ток: 
43. Связь между дискретным изменением тока через одиночные натриевые каналы с непрерывным изменением натриевого тока через мембрану (ансамбль каналов).
44. Ионные насосы в биологических мембранах: их виды, схемы действия. Сопряжённые процессы в ионных насосах.
При работе Nа/К–АТФазы за счет энергии высвобождающейся при гидролизе каждой молекулы АТФ в клетку переносятся два иона калия и выкачиваются три иона натрия. Таким образом, создается повышенная (по сравнению с межклеточной средой) концентрация в клетке ионов калия и пониженная концентрация ионов натрия.
Са-насос обеспечивает активный перенос двух ионов кальция, а протонный насос – двух протонов на одну молекулу АТФ.
Примерами активного транспорта является функционирование таких ионных насосов как Н+-АТФаза, Na+,К+-АТФаза, Са2+-АТФаза, Н+-РРаза, Cl--АТФаза. В электрогенезе высших растений, по-видимому, наибольшее значение имеет Н+-насос, создающий на плазмалемме и тонопласте электрохимические градиенты ионов Н+. Высокие значения мембранного потенциала в клетках растений в первую очередь создаются за счет транспорта ионов Н+ из цитоплазмы в окружающую среду.
45. ???Механизм распространения потенциалов действия вдоль нервного волокна, локальные токи, сальтаторное распространение. Скорость распространения потенциалов действия по нервному волокну.
Распространение Н. и. Н. и. может распространяться вдоль волокна без затухания и с пост. скоростью. Необходимая для передачи сигнала энергия не поступает из единого центра, а черпается в каждой точке волокна. Есть два способа передачи Н. и.: непрерывный и скачкообразный, когда импульс движется от одного перехвата к другому, перепрыгивая через области миелиновой изоляции.
В случае немиелинизир. волокна распределение мембранного потенциала j(x, t)определяется ур-нием:

где С - ёмкость мембраны, приходящаяся на единицу длины волокна, R - сумма продольных (внутриклеточного и внеклеточного) сопротивлений на единицу длины волокна, I - ионный ток, протекающий через мембрану волокна единичной длины. Электрич. ток I является функционалом от потенциала j, к-рый зависит от времени t и координаты х. Вид функционала I специфичен для биологически возбудимой среды. Однако ур-ние (5), если отвлечься от вида I, имеет более общий характер и описывает многие физ. явления, напр. процесс горения. Поэтому передачу Н. и. уподобляют горению порохового шнура. Если в бегущем пламени процесс поджигания осуществляется за счёт теплопроводности, то в Н. и. возбуждение происходит при помощи т. н. локальных токов (рис. 5).

Ур-ния Ходжкина - Хаксли для распространения Н. и. решались численно. Полученные решения вместе с накопленными эксперим. данными показали, что распространение Н. и. не зависит от деталей процесса возбуждения.
46. Электротонический потенциал. Зависимость электротонического потенциала от координаты волокна, формула, график. Постоянная длины волокна.
Повышение мембранного потенциала – величина деполяризующего потенциала V, передаваемого от возбужденных участков вдоль мембраны, зависит от расстояния х по формуле: V(х)=V0ех/l , V0 – повышение мембранного потенциала в зоне возбуждения, х – расстояние от возбужденного участка; l - константа длины нервного волокна равная расстоянию, на котором деполяризующий потенциал уменьшится в е раз.
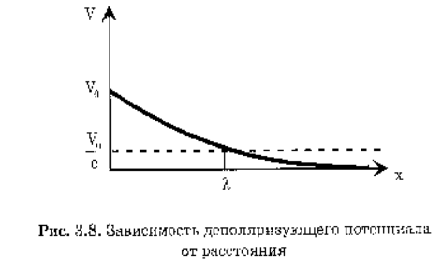 Константа длины нервного волокна:
Константа длины нервного волокна:  где rm – удельное электрическое сопротивление оболочки волокна, d - толщина оболочки, а - радиус нервного волокна, ri – удельное электрическое сопротивление цитоплазмы. Чем больше константа длины мембраны, тем больше скорость распространения нервного импульса. Величина l тем больше, чем больше радиус аксона и удельное сопротивление мембраны и чем меньше удельное сопротивление цитоплазмы. Так же увеличивают скорость миелиновые волокна.
где rm – удельное электрическое сопротивление оболочки волокна, d - толщина оболочки, а - радиус нервного волокна, ri – удельное электрическое сопротивление цитоплазмы. Чем больше константа длины мембраны, тем больше скорость распространения нервного импульса. Величина l тем больше, чем больше радиус аксона и удельное сопротивление мембраны и чем меньше удельное сопротивление цитоплазмы. Так же увеличивают скорость миелиновые волокна.
Дата добавления: 2015-02-10; просмотров: 1436; Мы поможем в написании вашей работы!; Нарушение авторских прав |