КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Жизнь в позднем палеозое
Как мы уже упоминали, начиная с верхнего карбона в Южном полушарии усилились процессы оледенения. На свободной от ледников территории Гондваны установился умеренный прохладный климат с выраженной сезонностью. Об этом говорит, в частности, наличие годичных колец в древесине гондванской флоры этого времени, получившей название глоссоптериевой по широко распространенным в ней видам семенного папоротника Glossopteris — небольшим растениям с крупными цельными листьями.
В состав глоссоптериевой флоры кроме различных семенных папоротников (птеридоспермов) входили также представители других групп голосеменных растений: кордаитов, гинкговых и хвойных. Глоссоптериевая флора была характерна для обширных территорий, соответствующих современным Индии, Афганистану, Южной Африке, Южной Америке, Австралии, Новой Зеландии и Антарктиде.
На северных континентах, входивших в состав Лавразии и располагавшихся в раннепермское время в значительной степени в экваториальном поясе, сохранялась растительность, близкая к тропической флоре карбона, но уже с меньшим богатством видов лепидодендронов и сигиллярий. К середине пермского периода во многих районах Лавразии климат становится все более засушливым, что приводит к изменению растительности: исчезают папоротники, каламиты, древовидные плаунообразные и другие влаголюбивые растения тропических лесов. Лишь на востоке Лавразии (современные Китай и Корея) климат и флора оставались в известной мере близкими к таковым в каменноугольное время.
Животный мир также претерпел значительные изменения на протяжении пермского периода, ставшие особенно драматичными во второй его половине. В наибольшей степени это коснулось морских животных, среди которых многие группы подверглись значительному угнетению, сопровождавшемуся уменьшением численности и разнообразия видов, вплоть до полного вымирания целых классов. Примерно в середине пермского периода вымирают последние трилобиты, эвриптериды, бластоидеи (один из классов прикрепленных иглокожих), палеозойские группы морских лилий. К концу перми исчезают тетракораллы, значительно сокращается число видов фораминифер среди простейших, морских ежей и офиур среди иглокожих, аммоноидей и наутилоидей среди моллюсков, остракод среди ракообразных, губок, мшанок и плеченогих. Из позвоночных вымирают акантодии, многие палеозойские группы хрящевых рыб. В пресных внутриконтинентальных водоемах значительно снижается численность хоановых рыб. К концу палеозоя вымирают лепоспондильные стегоцефалы. Изменения в наземной фауне были не столь значительны.
Пермское вымирание по своим масштабам принадлежит к категории так называемых «великих вымираний» и сравнимо, например, с широко известным великим вымиранием в конце мелового периода. Причины массовых вымираний, приводивших к радикальным изменениям общего облика фауны и флоры, во многом остаются загадочными. Вымирание вообще сопровождает эволюцию организмов как ее неизбежный побочный результат: в борьбе за существование выживают лишь те виды, которые оказались наиболее приспособленными при данных условиях. Вымирание отдельных видов происходило на любом отрезке эволюционной истории мира организмов либо в результате конкуренции с биологически близкими видами, либо по причине неспособности данного вида выдержать те или иные изменения абиотических факторов внешней среды. В обоих случаях возможности модифи-кационной изменчивости вымирающего вида и резерв его комбинативной изменчивости оказываются недостаточными, чтобы обеспечить его выживание в изменяющейся внешней среде.
В принципе те же общие причины определяют фатальную судьбу для каждого вымирающего вида и в эпохи массовых вымираний, но тогда одновременно обрывается множество филетических линий. Это может происходить и достаточно постепенно (численность вымирающих видов падает от поколения к поколению на протяжении больших промежутков времени), но в огромной исторической перспективе геологического времени эти процессы нередко выглядят как относительно быстрые и резкие изменения фауны и флоры.
Очевидно, массовые вымирания обусловлены какими-то причинами, имевшими общее значение для эволюционной судьбы сразу многих филетических линий, принадлежащих к разным группам организмов. Конечно, при этом совсем не обязательно, чтобы изменения какого-то одного или нескольких важных факторов прямо влияли на все подвергающиеся вымиранию виды организмов. Нарушения структуры биоценозов, выходящие за пределы возможностей естественной саморегуляции этих биологических макросистем, должны прямо или косвенно сказываться на многих видах организмов. Вероятно, существенной чертой эпох великих вымираний было именно разрушение многих биоценозов (биоценотические кризисы, по определению Б.Б.Родендорфа и В.В.Жерихина). Биоценотические кризисы могут быть вызваны самыми различными причинами, как абиотическими — изменения климата, рельефа и т. п., так и биотическими — появление и массовое расселение новых групп организмов, причем часто имеет место целый комплекс взаимодействующих факторов, что очень затрудняет анализ конкретных случаев, особенно в далеком прошлом.
О причинах пермского вымирания было выдвинуто несколько гипотез. Большинство из них в конечном итоге опирается на теорию дрейфа континентов. Согласно гипотезе, предложенной Д. Ва-лентайном и Э.Мурсом, образование Пангеи обусловило значительное сокращение общей площади мелководных бассейнов в области континентального шельфа, которая является наиболее благоприятной средой для жизни многих групп морских организмов. Это должно было привести к существенному нарушению многих сложившихся в середине палеозоя морских биоценозов и, как следствие, к вымиранию многих видов.
Другая гипотеза, выдвинутая Р. Боуэном, обращает внимание на потепление климата и завершение великого оледенения Гондваны во второй половине пермского периода, что могло быть связано с относительно небольшим смещением входивших в ее состав континентальных плит, в результате которого Южный полюс оказался в океане. Потепление климата привело к бурному таянию ледников и поступлению в океан огромного объема талых вод, которое должно было вызвать понижение солености морской воды. Пермское вымирание отразилось в первую очередь именно на тех группах морских организмов, которые наиболее чувствительны к изменениям солености: кораллы, морские лилии, мшанки, брахиоподы, головоногие моллюски и др. Новая стабилизация температурного и солевого режима океана произошла уже в триасовом периоде, и разнообразие форм морских беспозвоночных к концу триаса вновь стало возрастать.
Для наземных животных изменения физических условий в позднем палеозое не имели столь катастрофических последствий, как для представителей морской фауны. Пермский период стал временем расцвета древних пресмыкающихся, быстро осваивавших все новые местообитания, вытесняя хуже приспособленных конкурентов из числа стегоцефалов и батрахозавров, с запозданием пытавшихся закрепиться на суше.
Котилозавры разделились на несколько основных эволюционных стволов, первоначальная дифференциация которых была связана, вероятно, с освоением разных способов питания и различных местообитаний. Самыми крупными котилозаврами были растительноядные парейазавры (рис. 54), известные из средне- и верхнепермских отложений Европы и Африки. Эти животные, достигавшие 3 м длины, имели широкое массивное тело и крупный тяжелый череп с поразительно маленькой полостью, вмещавшей головной мозг. Наружная поверхность толстых черепных костей была покрыта своеобразным бугорчатым рельефом из приросших к ним остеодерм (вторичных кожных окостенений, часто встречающихся у рептилий), по бокам черепа выступали толстые костные пластины-«щеки» (отсюда название этих животных: Pareiasauria — «щекастые ящеры»). Парейазавры были, вероятно, неповоротливы и медлительны, от нападений хищников их защищал панцирь из толстых костных бляшек-остеодерм, рядами сидевших в коже. Судя по строению зубов (довольно слабых, с плоской листовидной коронкой), пищей парейазавров служили какие-то сочные и мягкие растения. Возможно, парейазавры обитали по берегам водоемов и болот, питаясь водной растительностью, и были палеозойскими аналогами современных бегемотов.

Рис. 54. Парейазавр Scutosaurus (реконструкция К.К.Флёрова)
Самые крупные хищники пермского времени принадлежали к подклассу зверообразных, или синапсидных рептилий (Theromorpha, или Synapsida). У этих пресмыкающихся в височной области черепа для его облегчения и предоставления необходимого пространства для сокращения мощных челюстных мышц возникла одна пара широких отверстий, или височных окон, пронизавших наружный черепной панцирь и ограниченных по бокам скуловыми дугами (рис. 55). Такой тип черепа, называемый синапсидным, дал одно из названий подкласса.

Рис. 55. Основные типы черепа амниот:
а — анапсидный; б — синапсидный; в — диапсидный; г — черепахи; д — ящерицы; е — птицы; 1 — височные окна; 2 — височные вырезки; 3 — задне-глазничная кость; 4 — чешуйчатая кость; 5 — скуловая кость; 6 — квадрат-носкуловая кость; 7 — квадратная кость. Стрелками указаны направления филогенетических преобразований
Самыми древними зверообразными рептилиями были пеликозавры (Pelycosauria), появившиеся уже в позднем карбоне. Их расцвет приходится на раннепермское время, когда многие их представители достигали крупных размеров, до 2 м. Некоторые пеликозавры (диметродон — Dimetrodon, эдафозавр — Edaphosaums и др.) имели странное приспособление: гипертрофированные остистые отростки спинных позвонков поднимались над позвоночником почти на 1 м, а у эдафозавра (рис. 56) на этих отростках имелись еще и короткие боковые выросты. Вероятно, между ними была натянута складка кожи, образуя огромный продольный гребень («спинной парус»). Может быть, эта складка играла роль в терморегуляции, увеличивая поверхность тела, что могло в одних ситуациях усиливать теплоотдачу, в других же — содействовать повышению температура тела при обогревании в солнечных лучах (повышение температуры тела путем инсоляции, называемое гелиотермией, присуще всем современным рептилиям).
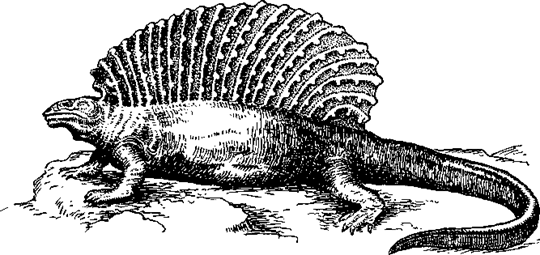
Рис. 56. Эдафозавр (Edaphosaurus) (реконструкция 3. Буриана)
Большинство пеликозавров вымерло уже в середине пермского периода, вероятно не выдержав конкуренции с представителями возникшей от пеликозавров более прогрессивной группы зверообразных рептилий — терапсид (Therapsida). В позднепермской эпохе терапсиды стали доминирующей группой пресмыкающихся. Этому способствовал целый ряд присущих им прогрессивных особенностей, в частности усовершенствование наземного передвижения. У большинства рептилий сохранилось примитивное положение конечностей, при котором плечо и бедро располагаются в целом горизонтально, локоть и колено направлены вбок, и тело животного удерживается на весу между конечностями усилиями мышц (отсюда характерное для рептилий «пресмыкание», когда брюхо при движении лишь незначительно приподнимается над землей). У высших терапсид локтевой сустав повернулся назад, коленный — вперед, бедро и плечо (и конечности в целом) оказались расположенными под телом, высоко приподнятым над субстратом; для поддержки тела при этом не требуется столь значительной работы мышц, как у типичных пресмыкающихся.
У многих терапсид произошла дифференциация зубной системы на передние острые «резцы», крупные кинжалообразные «клыки» и заклыковые щечные зубы, которые у высших представителей этой группы (териодонтов, или «зверозубых», — Theriodontia) приобрели усложненную коронку с тремя бугорками, что позволяло более эффективно пережевывать пищу. Пережевывание пищи в ротовой полости требовало задержки дыхания, в связи с чем у терапсид развивается вторичное нёбо, разделившее примитивную ротовую полость на два этажа: верхний (носовой ход) и нижний (собственно ротовая полость).
Терапсиды были очень разнообразны: среди них были и хищники различных размеров, вплоть до трехметровых иностранцевий (Inostrancevia, рис. 57) с клыками более 10 см длиной (своего рода «саблезубых тигров» пермского периода), и растительноядные животные, иногда имевшие причудливое строение.

Рис. 57. Иностранцевия (Inostrancevia) (реконструкция К.К.Флёрова)
Для некоторых растительноядных дейноцефалов (Deinocephalia, их название буквально означает «страшноголовые») характерны большие костные бугры или шишки на темени и щечной области черепа, придававшие голове этих животных устрашающий вид (скорее всего, это были защитные приспособления). Были и хищные дейноцефалы, например приземистый и длинный титанофонеус — Titanophoneus, большие лапы которого, по мнению Ю.А. Орлова, были снабжены плавательными перепонками, а сам этот хищник вел полуводный образ жизни.
В позднепермское время были широко распространены дицинодонты (Dicynodontia), утратившие все зубы, кроме пары огромных верхних клыков, сохранявшихся у некоторых видов лишь у самцов. Беззубые челюсти были покрыты роговым клювом. Нелегко представить себе способ питания этих странных животных, среди которых были и сравнительно мелкие, и очень крупные, размеров носорога, виды. Предполагали, что дицинодонты питались падалью или поедали питательную сердцевину стволов саговниковых растений, раскалывая или разрывая их своими клыками, или выкапывали корневища хвощеобразных растений, в изобилии произраставших по берегам водоемов.
Местонахождения с ископаемыми остатками целых комплексов видов позднепермских рептилий (котилозавров, дицинодонтов, дейноцефалов, териодонтов) с довольно сходным общим обликом фауны известны в Южной Африке и Восточной Европе. Замечательные местонахождения позднепермских тетрапод были обнаружены на территории России. На берегу Северной Двины, в районе г. Котласа, В. П. Амалицким в 1897 г. были найдены остатки крупных парейазавров, дицинодонтов, хищных териодонтов (среди них были огромные иностранцевии), батрахозавров и лабиринтодонтов. Еще одна интересная позднепермская фауна была раскопана в 1957—1960 гг. в Очёрском районе Пермской области. В состав Очёрской фауны, исследованной П.К.Чудиновым, входили хищные и травоядные дейноцефалы (среди последних огромные рогатые эстемменозухи, Estemrnenosuchus), хищные териодонты, терапсиды, близкие к предкам дицинодонтов, лабиринтодонты. Костеносные породы этих местонахождений сформировались из глинистых и песчаных осадков речных дельт[50].

Рис. 58. Никтифрурет (Nyctiphraretus) (реконструкция А. П. Быстрова)
В некоторых филогенетических стволах рептилий продолжалось совершенствование приспособлений к насекомоядности, которая была, вероятно, самой примитивной формой питания пресмыкающихся. Это направление эволюционных преобразований привело к появлению небольших и проворных ящерицеобразных животных, причем сходный тип строения сформировался параллельно в нескольких независимых фйлетических линиях. Одна из них сохранила общий уровень организации котилозавров, со сплошным покровом накладных костей в височной области черепа. Это были проколофоны (Procolophonia). Остатки примитивных проколо-фонов, никтифрурета (Nyctiphruretus, рис. 58) и никтеролетера (Nycteroleter) — легких большеглазых животных с удлиненным телом и хвостом общей длиной до 30—40 см были найдены в верхнепермских отложениях на берегах р. Мезени.
В других группах насекомоядных рептилий произошли некоторые прогрессивные изменения организации, в частности усовершенствование строения черепа. Эти животные, давшие начало двум центральным подклассам рептилий: лепидозаврам (Lepidosauria) и архозаврам (Archosauria) — приобрели удлиненные челюсти, удобные для схватывания мелкой подвижной добычи, и две пары височных окон, расположенных друг над другом и разделенных тонким костным мостиком — верхней височной дугой. Так возник диапсидный тип черепа (см. рис. 55, в), легкая и ажурная конструкция которого позволила усовершенствовать так называемый кинетизм — подвижность верхней челюсти и некоторых связанных с ней костей, унаследованную наземными позвоночными от кистеперых рыб (рис. 59).
Рис. 59. Кинетизм черепа ящерицы-игуаны Cyclura.
Заштрихован неподвижный затылочный сегмент черепа (задняя часть мозговой коробки) сплошным контуром и штриховой линией показаны два положения движений максиллярного сегмента (верхней челюсти и связанных с ней костей черепа)
Примитивные лепидозавры (миллерозавры — Millerosauria), у которых лишь началось развитие височных окон и усиление кинетизма черепа, известны из верхнепермских отложений Южной Африки. В породах примерно того же возраста сохранились остатки первых рептилий, имевших настоящий диапсидный череп — эозухий (Eosuchia), наиболее известным представителем которых была янгина (Youngina). Отдаленным потомком эозухий является дожившая до нашего времени новозеландская гаттерия (Sphenodon), которая принадлежит к отряду клювоголовых (Rhynchocephalia), появившихся в раннем триасе. От эозухий или от миллерозавров возникли настоящие ящерицы (Lacertilia), у которых в связи с усилением подвижности верхней челюсти (см. рис. 59) редуцировалась нижняя височная дуга и череп приобрел легкую ажурную конструкцию. Ящерицы достоверно известны начиная со среднего триаса.
Вероятно, эозухий были также предками высших рептилий, принадлежащих к подклассу архозавров, которые оставили самые яркие страницы в палеонтологической истории мезозоя. Д.Уотсон высказал предположение, что обособление предков архозавров от ранних насекомоядных лепидозавров было связано с переходом первых к питанию позвоночными. Этим объясняется более массивная конструкция черепа архозавров и развитие у них более крупных зубов, сидящих в отдельных ячейках (текодонтность). Для архозавров характерно также более выпрямленное положение конечностей, чем у лепидозавров.
Наиболее ранним известным архозавром является позднепермский проторозавр (Protorosauras), пропорциями тела и размерами (1—2 м) напоминавший современных ящериц-варанов.
Дата добавления: 2015-04-16; просмотров: 351; Мы поможем в написании вашей работы!; Нарушение авторских прав |