КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Освоение суши
Первые вполне достоверные остатки наземных организмов известны начиная с силура[46]. Заселение безжизненных прежде материков было длительным процессом, развивавшимся в течение нескольких десятков миллионов лет, на протяжении ордовика, силура и девона.
Вероятно, начало освоению суши положило появление пленчатых обрастаний прибрежных камней и скал в полосе прибоя и приливно-отливной зоне синезелеными водорослями; в водорослевых обрастаниях поселялись бактерии. В условиях низких континентов раннего палеозоя с очень пологими берегами эта переходная зона между морем и сушей имела значительную протяженность. Сформировавшиеся здесь пленчатые бактериально-водорослевые обрастания («маты») часто перекрывались неорганическими материалами — продуктами выветривания горных пород, сносившимися с суши. В результате этих процессов происходило первичное почвообразование. Формирование почвы стало предпосылкой для освоения суши растениями и животными, что, в свою очередь, усилило процессы почвообразования. Этому способствовало накопление органических веществ. Внесение в почву лигнина — химически устойчивого углевода, содержащегося в ксилеме сосудистых растений, замедляло процессы разложения почвенной органики. Почва аэрировалась ходами, которые проделывали в ней почвенные животные (черви и членистоногие).
Древнейшими растениями, имевшими в основном «наземный» облик, но населявшими, вероятно, пограничную зону между водой и сушей и влажные местообитания в низменных прибрежных районах, были псилофиты (Psilophyta, или Rhyniophyta) из группы риниевых (Rhyniales) — невысокие травянистые или кустар-никовообразные растения (Rhynia, Cooksonia), еще не имевшие настоящих корней и листьев; органы бесполого размножения (спорангии) располагались на концах дихотомически разветвленных побегов (рис. 46).

Рис. 46. Псилофиты (реконструкция из А.Н.Криштофовича, 1957):
а — Rhynia; б — Asteroxylon
В их древесине уже имелись примитивные сосудистые образования — трахеиды. Вероятно, в жизненном цикле этих растений происходила четкая смена поколений — диплоидного спорофита и более просто устроенного гаплоидного гаметофита.
Риниевые могли возникнуть от каких-то высших водорослей (может быть, близких к харовым) еще в ордовике. Псилофиты ныне рассматриваются как сборная группа, объединяющая древнейшие наземные растения — риниевые и возникшие от них зостерофиллофиты (Zosterophyllales) и тримерофиты (Trimero-phytales), обладавшие более развитым ветвлением с особыми спо-рангиеносными ветвями, мелкими листочками и некоторыми другими прогрессивными особенностями. Псилофиты известны начиная со среднего силура и достигли расцвета в раннем и среднем девоне.
В верхнесилурийских отложениях встречаются также ископаемые остатки спор и гиф, которые интерпретируют как древнейшие известные остатки наземных грибов — аскомицетов. Вероятно, грибы принимали важное участие в процессе почвообразования.
К этому времени (силур — девон) относится и возникновение наиболее древних толщ горючих ископаемых, образующихся при неполном разложении скоплений растительных остатков.
Вероятно, в самом раннем девоне или даже в позднем силуре от самых примитивных риниевых могли возникнуть мохообразные (Bryophyta), а от высших псилофитов произошли различные группы сосудистых растений, приобревшие листья, корневую систему и более совершенные способы размножения: от зостерофиллофитов — плаунообразные (Lycopsida), а от тримерофитов — членистостебельные, или хвощевые (Sphenopsida) и папоротникообразные (Pteropsida). Представители этих групп растений, лучше приспособленных к жизни на суше, в позднем девоне повсеместно вытеснили псилофитов и сформировали первую настоящую наземную флору, включавшую и древовидные формы. Эту флору называют «археоптерисовой» по широко представленным в ней видам папоротников Archaeopteris. К этому времени относится и появление первых голосеменных (Gymnospermae), называемых птеридоспермами или семенными папоротниками — Pterido-spermae, возникших от древних разноспоровых папоротников и еще сохранявших внешний облик древовидных папоротникообразных.
В процессе приспособления к жизни на суше у растений совершенствовались вегетативные органы, в которых дифференцировались различные ткани: покровные (защищавшие растение от избыточного испарения и различных повреждений), механические (опорные, особенно важные в условиях возросшей в воздушной среде силы тяжести) и проводящие (обеспечивавшие транспорт веществ между различными органами). Особенно сложной для растений на суше стала проблема осуществления полового процесса: в воде подвижные мужские гаметы легко достигали яйцеклеток, тогда как в воздушной среде при неподвижности растений встреча гамет невозможна без специальных приспособлений, обеспечивающих их перенос от одного растения к другому. У высших наземных растений такие приспособления сформировались из гаплоидного поколения (гаметофита), которое редуцируется до одной — нескольких клеток (пыльца семенных растений). Диплоидное же поколение (спорофит) становится основным в жизненном цикле. Противоположное соотношение этих поколений в жизненном цикле — с преобладанием гаметофита — имеется лишь у мхов, что, вероятно, решающим образом ограничило адаптивные возможности этой группы в освоении суши: мхи выживают лишь в условиях достаточно высокой влажности и тесно связаны с субстратом.
Во влажном и теплом климате, характерном для первой половины каменноугольного периода, широкое распространение получила обильная наземная флора, имевшая характер густых влажных тропических лесов (рис. 47).

Рис. 47. Лесной ландшафт карбона (по З.Буриану).
Слева на переднем плане каламиты, за ними — сигиллярии, правее на переднем плане — семенной папоротник, вдали в центре — древовидный папоротник, справа — лепидодендроны и кордаиты
Среди древовидных растений выделялись представители плаунообразных — лепидодендроны (Lepidodendraceae), достигавшие 40 м в высоту и до 2 м в диаметре у основания, и сигиллярии (Sigillariaceae) тридцатиметровой высоты. Стволы этих деревьев были покрыты характерным рельефным узором, образованным следами отпавших листьев (у взрослых деревьев листья сохранялись лишь в высокой кроне, где ствол дихотомически ветвился). Среди этих гигантов палеозойского леса высились и хвощеобразные пирамидальные каламиты (Calamitales) с членистым стеблем и мутовками мелких жестких листьев. Очень обильны были различные папоротники, как ползучие, так и древовидные, хотя и не достигавшие такой высоты, как плаунообразные, и составлявшие второй ярус лесной растительности и подлесок. Голосеменные были представлены разнообразными птеридоспермами, а также новой группой — кордаитами (Cordaitales). Кордаиты были высокими деревьями, стволы которых напоминали знакомые нам хвойные, но высоко расположенные кроны были образованы крупными, до 1 м в длину, ланцетообразными или лентовидными листьями. В древесине всех этих деревьев не обнаружено годичных колец, что говорит об отсутствии четко выраженной сезонности климата.
Наличие густых тропических лесов вдоль морских побережий, изрезанных лагунами, пересеченных дельтами рек и на больших территориях заболоченных, создавало предпосылки для интенсивного углеобразования. Мощные толщи каменных углей, сформировавшиеся в это время в разных районах земного шара, дали название каменноугольному периоду.
По мере заселения суши растениями возникли предпосылки для освоения наземной среды обитания животными. Вероятно, первыми на суше появились какие-то мелкие беспозвоночные, питавшиеся органическими остатками в почве. Возможно, такие животные появились уже к середине силурийского периода. Древнейшие известные наземные членистоногие — мелкие плоскоспинные многоножки (Myriapoda) со сдвоенными сегментами тела (как у современных двупарноногих — кивсяков) — были обнаружены в позднесилурийских и раннедевонских отложениях. М. С. Гиляров показал, что для многих групп беспозвоночных животных освоение суши началось с использования почвы как промежуточной среды обитания, в которой ряд факторов был в какой-то степени близок к показателям водной среды (высокая влажность, плотность субстрата). В почве можно использовать многие приспособления, сложившиеся в воде, при относительно незначительной их перестройке (аппарат передвижения, органы дыхания и др.). Вероятно, этим путем на сушу проникли из водоемов различные группы червей, которые в основном и остались почвенными обитателями, и членистоногих. Жизнь в почве и близких к ней местообитаниях характерна для наиболее примитивных современных групп членистоногих (онихофоры, многоножки, низшие насекомые — аптериготы и др.).
Приспособление животных к жизни на суше требовало, во-первых, перестройки локомоторного аппарата для передвижения в условиях возросшей по сравнению с водой силы тяжести; во-вторых, развития органов дыхания, способных усваивать кислород воздуха и избавлять организм от углекислого газа; в-третьих, формирования приспособлений, защищающих организм от обезвоживания. Последние включают развитие кожных покровов, снижающих потери воды с испарением (эпикутикула у членистоногих), а также совершенствование органов выделения, которые должны выводить из организма экскреты в малотоксичной и концентрированной (обезвоженной) форме. У водных животных чаще всего конечным продуктом азотистого обмена является аммиак, ядовитый и требующий для своего растворения значительного количества воды. У наземных форм выводятся из организма такие соединения, как мочевина, мочевая кислота, гуанин и др.
Замечательно, что очень сходные приспособления к наземной жизни формировались независимо и параллельно у представителей разных эволюционных стволов: в типе членистоногих, например, у нескольких самостоятельных линий среди хелицеровых (у скорпионов, пауков, сольпуг, клещей); во многом сходные адаптации развивались и у трахейных (у многоножек и насекомых). Отметим, что указанные выше приспособления растений к на-земности также формировались независимо и параллельно у плаунообразных, членистостебельных, папоротникообразных. Параллельная эволюция вообще очень широко распространена в природе. Причины этой закономерности будут рассмотрены в последней части книги.
Из девона известны представители нескольких групп наземных членистоногих: палеозойская группа панцирных пауков (Soluta), клещи (Acaromorpha) и низшие первичнобескрылые группы насекомых (Insecta). Несомненно, разнообразие наземных беспозвоночных в девонском периоде было значительно большим, чем это представляется по дошедшим до нас скудным ископаемым остаткам.
Во второй половине раннекарбоновой эпохи появились наделенные крыльями высшие насекомые — птериготы. Вероятно, предки крылатых насекомых перешли к лазающей жизни на стволах и в кронах деревьев. Для планирования при падениях и прыжках у этих животных служили боковые уплощенные выступы тела на грудных сегментах. Из этих неподвижных придатков в процессе приспособительной эволюции естественный отбор сформировал удивительный в своем совершенстве летательный аппарат. Первоначально крылья имелись на всех 3 сегментах груди, хотя передняя пара еще сохраняла примитивное строение в виде неподвижных плоскостей, как это было, например, у палио-диктиоптеры (Palaeodictyoptera) (рис. 48).

Рис. 48. Палеодиктиоптера (Palaeodictyoptera)
У большинства известных групп насекомых эта передняя пара крыльев утрачена. К концу карбона уже существовали разнообразные группы летающих насекомых, некоторые из них дожили до современности (поденки, стрекозы, тараканы, прямокрылые, равнокрылые, скорпионницы). В палеозое насекомые были единственными летающими животными. Некоторые виды стрекоз тогда достигали крупных размеров: у верхнекарбоновых — нижнепермских Meganeura длина тела достигала 30 см, размах крыльев до 65 см.
Разнообразие наземных беспозвоночных в карбоне было дополнено появлением растительноядных брюхоногих моллюсков (Gastropoda) из группы легочных (Pulmonata), дышащих воздухом. Вслед за беспозвоночными к освоению наземной среды обитания приступили и позвоночные. Их выход на сушу был возможен лишь после появления соответствующей пищевой базы (достаточно обильной фауны наземных беспозвоночных). Эти крупные животные не могли использовать почву как промежуточную среду между водой и сушей. Поэтому освоение суши для позвоночных было возможно только в условиях влажного и теплого климата, наиболее благоприятного для животных, физиология которых еще во многом подобна физиологии рыб. С другой стороны, комплекс преадаптаций для перехода к наземной жизни должен был сформироваться у предков наземных позвоночных еще в водной среде.
Предками наземных позвоночных (тетрапод) стали кистеперые рыбы, обладавшие легкими и снабженными мощной мускулатурой парными конечностями — саркоптеригиями, которые, вероятно, позволяли этим рыбам не только передвигаться по дну водоемов среди густых зарослей, но и выбираться на мелководье, проползать по мелким протокам из одного водоема в другой. Внутренний скелет этих конечностей был образован одной крупной костью, сочлененной с поясом конечностей (плечевым или тазовым), далее следовал второй отдел из двух параллельных костей, и дистально располагались многочисленные мелкие косточки (рис. 45, в). В этой схеме легко просматривается прототип скелета конечностей наземных позвоночных.
Вероятно, некоторые виды кистеперых рыб перешли к освоению новой кормовой базы — обильной фауны беспозвоночных, обитавших на мелководье, среди прибрежных растений и по берегам водоемов. Это направление приспособительной эволюции вело дальше к освоению суши, с ее бурно развивавшейся флорой и фауной, в составе которой еще не было крупных хищников. Эту экологическую нишу и заняли наземные позвоночные.
Отметим, что приспособления к наземности независимо и параллельно развивались в нескольких линиях эволюции кистеперых рыб. В связи с этим Э.Ярвик выдвинул гипотезу о дифилети-ческом происхождении наземных позвоночных от двух разных групп кистеперых рыб (Osteolepiformes и Porolepiformes). Однако целый ряд ученых (А.Ромер, И.И.Шмальгаузен, К.Томсон, Э.И.Воробьева) подвергли аргументы Ярвика критике. Ныне большинство исследователей считают более вероятным монофилетическое происхождение тетрапод от остеолепиформных кистеперых, хотя при этом допускается возможность парафилии, т. е. достижения уровня организации земноводных несколькими близкородственными филетическими линиями остеолепиформных рыб, эволюционировавшими параллельно.
Наиболее древние представители земноводных (класс Amphibia) ихтиостеги (Ichthyostegalia, рис. 49) известны из верхнедевонских отложений Гренландии. Ихтиостеги были довольно крупными животными (свыше 80 см длиной), имевшими уже хорошо развитые пятипалые конечности наземного типа, с помощью которых они, несомненно, могли переползать по суше[47]. Однако значительную часть жизни ихтиостеги, вероятно, проводили в водоемах, будучи, по удачному выражению Ярвика, «четвероногими рыбами». Об этом говорит наличие у них настоящего хвостового плавника, напоминавшего хвост кистеперых рыб, а также сохранение во взрослом состоянии сейсмосенсорных рецепторов («боковой линии» — органов чувств, которые функционируют только в водной среде, воспринимая инфразвуковые колебания и электромагнитные поля). В черепе ихтиостег сохранялись рудименты костей жаберной крышки (возможно, имелись и жабры, укрепленные на хрящевых жаберных дугах). Тело было покрыто мелкими костными чешуйками.


Рис. 49. Ихтиостега (Ichthyostega) (по Э.Ярвику)
Вероятно, древнейшие земноводные, подобные ихтиостегам, в водной среде проигрывали конкуренцию многочисленным и разнообразным кистеперым рыбам, поскольку парные конечности наземного типа не могли быть эффективными рулями глубины при плавании. Преимущества пятипалых конечностей проявлялись лишь в наиболее мелких участках водоемов и на суше. Скорее всего, между кистеперыми рыбами и их потомками — земноводными—в девоне существовали конкурентные отношения, способствовавшие расхождению этих групп в разные адаптивные зоны: кистеперые рыбы доминировали в водоемах, а земноводные заняли промежуточные местообитания между водой и сушей (мелкие прибрежные участки водоемов, где свободное плавание затруднено; заболоченные районы; области с избыточной влажностью на суше). Попутно отметим, что этот тип местообитаний, промежуточных и пограничных между водой и сушей, остался и доныне наиболее благоприятной для земноводных адаптивной зоной, где амфибии сохранили преимущества и перед рыбами, и перед более совершенными наземными группами тетрапод. В позднем девоне уже началась дивергентная эволюция древнейших земноводных. Помимо ихтиостег в конце девона существовали представители и ряда других групп амфибий, ископаемые остатки которых были обнаружены в различных районах. Любопытно, что число пальцев у этих животных часто достигало 7—8.
В карбоне начинается расцвет древних амфибий, представленных в верхнем палеозое большим разнообразием форм (рис. 50), которых условно объединяют под названием стегоцефалов, т. е. покрытоголовых. Этот термин, не имеющий ныне таксономического значения, подчеркивает характерную особенность этих животных, голова которых была защищена унаследованным от рыб сплошным костным панцирем (так называемый стегальный череп), пронизанным лишь отверстиями ноздрей, глазниц и «теменного глаза».

Рис. 50. Представители стегоцефалов:
а — Pteroplax (Eogyrinus); б — Eryops; в — Cacops (реконструкции А. Панчена, А. Ромера, Е. Олсона)
Наиболее известной из групп стегоцефалов являются лабиринтодонты (Labyrinthodontia), к числу которых принадлежат и ихтиостеги. Их название связано с любопытной особенностью строения зубов, также унаследованной от кистеперых рыб: эмаль и дентин образуют многочисленные складки, так что на поперечном шлифе возникает картина, напоминающая лабиринт.
Лабиринтодонты в верхнем палеозое были одной из наиболее распространенных и обильных видами групп позвоночных. К ним принадлежали и мелкие, и крупные, более 1,5 м, формы. В карбоне преобладали виды со слабыми конечностями и длинным телом, которые, вероятно, обитали в многочисленных болотах. В пермское время появились крупные крокодилообразные стегоцефалы с большой уплощенной головой (подобные Eryops, рис. 50, б), а также мелкие виды с лучше развитыми конечностями и укороченным туловищем и хвостом (Cacops, рис. 50, в). Эти последние жили, скорее всего, преимущественно на суше, питаясь различными беспозвоночными, хотя, как и все земноводные, нуждались в повышенной влажности воздуха и близости водоемов для откладки икры. Как считает ряд ученых, мелкие лабиринтодонты, державшиеся по берегам водоемов и спасавшиеся от наземных врагов прыжками в воду, стали предками бесхвостых земноводных (Anura), возникших, вероятно, в позднепермское время. Древнейший известный представитель бесхвостых амфибий Triadoba-trachus из раннего триаса Мадагаскара уже имел характерные пропорции тела и основные структурные особенности лягушек, хотя еще сохранял небольшой хвост.
Два других отряда современных амфибий в ископаемом состоянии известны из значительно более поздних отложений: хвостатые (Urodela) из средней юры, безногие, или червяги (Apoda), из ранней юры. Проблема их происхождения остается дискуссионной. По многим признакам скелета и мускулатуры головы хвостатые амфибии близки к бесхвостым и, возможно, имеют с ними общее происхождение.
Безногие амфибии стоят несколько особняком, сохраняя во многих отношениях наиболее примитивное строение среди современных земноводных. Некоторые палеонтологи связывают их происхождение с филогенетическим стволом стегоцефалов, объединяемых в подкласс лепоспондильных (Lepospondyli)[48].
Среди лепоспондильных было много форм с сильно удлиненным телом и слабо развитыми конечностями. Таковы, например, лизорофы (Lysorophia). Представители другого отряда лепоспондильных — аистоподы (Aistopoda), как и червяги, совсем утратили конечности. Однако они не могли быть предками червяг, так как обладали рядом специфических особенностей строения. К ле-поспондильным принадлежат также нектридии (Nectridia). Одно из их семейств отличалось причудливым строением черепа, задние углы которого непомерно разрастались вбок и назад в виде длинных конических выростов, так что голова в целом сверху выглядела подобной бумерангу. Предполагают, что эти выросты могли играть роль своего рода подводных крыльев, создававших при движении некоторую подъемную силу, способствуя планированию в воде.
От примитивных лабиринтодонтов уже в девоне обособилась группа антракозавров (Anthracosauria), древнейший представитель которых Tulerpeton был обнаружен в верхнедевонских отложениях Тульской области. Первоначально эти стегоцефалы также обитали в прибрежной зоне водоемов. Однако среди антракозавров очень рано наметилась тенденция к освоению наземных местообитаний. Как и в большинстве случаев, эта тенденция независимо и параллельно развивалась в нескольких разных филетических линиях, в каждой из которых постепенно формировались во многом сходные приспособления к жизни на суше. Антракозавров и тех из их потомков, которые сохранили, несмотря на наземный образ жизни, общий уровень организации и характерные черты размножения и онтогенеза земноводных (с откладкой икры в водоемах и водными личинками, имеющими жабры), объединяют в подкласс батрахозавров, или «ящероземноводных» (Batrachosauria). К батрахозаврам принадлежат сеймурии (Seymouria; рис. 51), котлассии (Kotlassia) и родственные им формы, существовавшие в пермском периоде. Сеймурии долгое время считали не земноводными, а пресмыкающимися, так близки они по строению черепа, позвоночника и конечностей к настоящим рептилиям. Однако позднее были обнаружены остатки личиночных стадий этих животных, имевших следы каналов сейсмосенсорных органов на костях черепа и, следовательно, живших в воде.
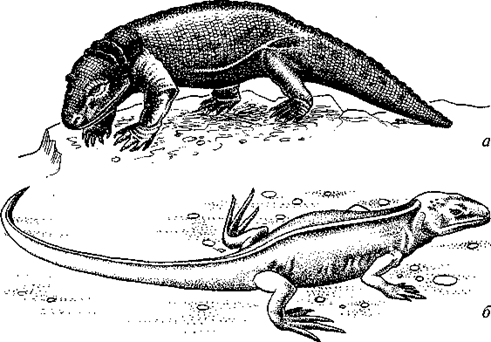
Рис. 51. Батрахозавры:
а — сеймурия (Seymouria); б — гефиростегус (Gephyrostegus) (реконструкции
З.Буриана и Д.Уотсона)
От каких-то примитивных батрахозавров в раннем карбоне возникли настоящие рептилии (Reptilia), к которым, в частности, относят небольшую форму Westlothiana, обнаруженную в нижне-карбоновых отложениях Шотландии. В ее организации сочетаются прогрессивные (рептильные) особенности черепа и примитивное состояние ряда признаков позвоночника и конечностей («моза-ичность»). Р. Кэррол считает, что непосредственные предки пресмыкающихся могли быть близки по основным особенностям к позднекарбоновым гефиростегам (Gephyrostegidae, рис. 51), которые, однако существовали слишком поздно, чтобы быть действительными предками рептилий. Возможно, гефиростеги были реликтовой группой, сохранившей ряд архаических особенностей строения, характерных для раннекарбоновых предков пресмыкающихся.
Несомненные рептилии известны начиная со среднего карбона. Е.Олсон указал на близкое совпадение времени появления пресмыкающихся и разнообразных групп высших насекомых-птеригот. Отметив, что все наиболее древние рептилии были относительно мелкими насекомоядными животными, Олсон пришел к выводу, что формирование пресмыкающихся как группы было тесно связано с использованием новой богатой пищевой базы, возникшей на суше с появлением насекомых.
Вероятно, уже в карбоне у пресмыкающихся сформировался целый комплекс приспособлений, позволивших им и их потомкам стать в полной мере наземными животными. Важнейшим из этих приспособлений, которое сделало возможными многие другие существенные изменения организации пресмыкающихся, было развитие эффективного механизма вентиляции легких посредством движений грудной клетки. Он сделал ненужным кожное дыхание, которое у земноводных является необходимым дополнением легочного, так как из-за несовершенства вентиляции легких, где скапливается избыток углекислого газа, земноводные нуждаются в выведении последнего через кожу. Поэтому в коже рептилий стало возможным усиление процессов ороговения эпидермиса, защитившего организм от постоянной потери влаги через покровы (неизбежного при кожном дыхании у земноводных) и риска гибели в сухом воздухе от обезвоживания. После этого пресмыкающиеся получили возможность заселить практически любые местообитания на суше, вплоть до безводных пустынь. Этому способствовало также развитие метанефрической (тазовой) почки и механизмов обратного всасывания воды из мочи, позволявших выводить из организма максимально концентрированные экскреты. После ликвидации кожного дыхания у рептилий стало возможным разделение в сердце артериальной (поступающей из легких) и венозной (идущей от всех остальных органов) крови, что также невозможно для земноводных, так как у них от кожи кровь поступает в правое предсердие, а от легких — в левое, и разделение этих потоков крови в желудочке сердца затруднило бы использование кожного дыхания. Все эти важнейшие достижения рептилий наряду с целым рядом усовершенствований скелета (увеличение подвижности головы с развитием особой формы передних шейных позвонков, некоторые преобразования в скелете конечностей и др.) создали важнейшие предпосылки для полного овладения наземной средой обитания.
Одновременно у древнейших рептилий сформировался второй комплекс приспособлений, избавивших от связи с водоемами все процессы, связанные с размножением. Это было появление внутреннего осеменения вместо характерного для земноводных наружного; формирование крупных яиц, обильно снабженных запасами питательных веществ (желтком) и способных обеспечить длительное развитие зародыша; образование прочных яйцевых оболочек, защищающих яйцо на суше от обезвоживания и механических повреждений; и наконец, возникновение особых зародышевых оболочек — амниона, серозы и аллантоиса, обеспечивающих благоприятную среду для развития зародыша, газообмен и накопление экскретов (из-за невозможности их выведения из яйца во внешнюю среду). По признаку наличия этих зародышевых оболочек высшие классы наземных позвоночных объединяются в группу амниот, противопоставляемую низшим позвоночным — анамниям.
Древнейшие и наиболее примитивные пресмыкающиеся принадлежали к подклассу котилозавров (Cotylosauria)[49]. Название «котилозавр» подчеркивает характерную особенность черепа этих животных, защищающего голову сплошным костным панцирем, как у стегоцефалов, но более высокого и узкого и несколько напоминающего у некоторых котилозавров при взгляде сверху чашу или кубок, откуда и название (греч. kotylos — чаша, кубок). Центральной группой котилозавров являются капториноморфы (Captor-hinomorpha, рис. 52), древнейшие представители которых известны из среднекарбоновых отложений. Это были небольшие, до 50 см длиной, насекомоядные животные.
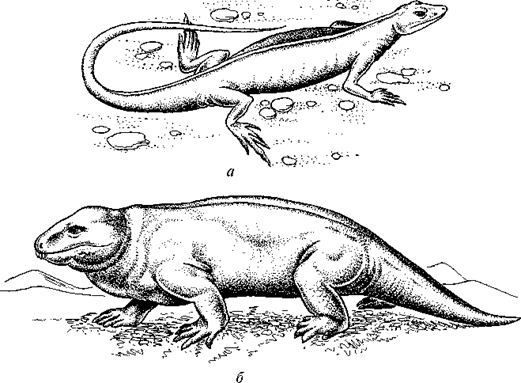
Рис. 52. Представители котилозавров:
а — капториноморф Hylonomus (по Р.Кэрролу); б — диадект Diadectes (по Е.Олсону)
Уже во второй половине каменноугольного периода возникло большое разнообразие форм пресмыкающихся, среди которых появились помимо насекомоядных растительноядные животные (диадектиды) и крупные хищники, питавшиеся позвоночными (некоторые более поздние капториноморфы и другие группы). Удивительно, что некоторые филетические линии рептилий уже в карбоне вернулись в водоемы, став полуводными или даже полностью водными животными. Особенно интересны мезозавры (Mesosauria, рис. 53), конечности которых преобразовались в ласты, а длинные узкие челюсти (удобные для схватывания добычи в воде) были усажены многочисленными тонкими и острыми зубами. Остатки мезозавров были найдены в Южной Африке и Бразилии. Это хорошо соответствует представлениям о существовании в верхнем палеозое единого материка Гондваны.

Рис. 53. Мезозавр (Mesosaurus) (по З.Буриану)
Дата добавления: 2015-04-16; просмотров: 534; Мы поможем в написании вашей работы!; Нарушение авторских прав |