КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Л Е К Ц І Ї
1. Розглянемо поверхню другого порядку  , задану рівнянням
, задану рівнянням
 , (1)
, (1)
де  - деякі дійсні числові коефіцієнти, причому коефіцієнти
- деякі дійсні числові коефіцієнти, причому коефіцієнти  одночасно не дорівнюють нулю.
одночасно не дорівнюють нулю.
Доданки  називають групою старших членів, або квадратичною формою, вираз
називають групою старших членів, або квадратичною формою, вираз  - лінійною частиною, число
- лінійною частиною, число  - вільним членом рівняння.
- вільним членом рівняння.
Рівняння (1) називають загальним рівнянням поверхні другого порядку, оскільки з нього можна отримати будь-яке конкретне рівняння другого порядку. З поверхнями, рівняння яких є частинними випадками рівняння (1), ми уже зустрічалися в попередніх лекціях. Такими є сфера  із центром у точці
із центром у точці  із радіусом
із радіусом  , еліпсоїд
, еліпсоїд  , одно- та двопорожнинні гіперболоїди
, одно- та двопорожнинні гіперболоїди  , еліптичний та гіперболічний параболоїди
, еліптичний та гіперболічний параболоїди 
 та інші. Природно виникає питання, чи всі можливі поверхні другого порядку ми розглянули? Приклад поверхні, яка задається рівнянням
та інші. Природно виникає питання, чи всі можливі поверхні другого порядку ми розглянули? Приклад поверхні, яка задається рівнянням  , показує, що ні. Справді, записавши дане рівняння у виді
, показує, що ні. Справді, записавши дане рівняння у виді  , бачимо, що його не задовольняють координати жодної точки. Можна навести також інші приклади рівнянь поверхонь, які ми ще не розглядали.
, бачимо, що його не задовольняють координати жодної точки. Можна навести також інші приклади рівнянь поверхонь, які ми ще не розглядали.
Оскільки рівняння (1) містить 10 коефіцієнтів, які визначаються з точністю до сталого множника, то поверхня другого порядку задається не більше, ніж 9-ма точками. В деяких випадках їх кількість може бути меншою. Наприклад, сферу можна задати 4-ма точками, які їй належать. Меншою кількістю точок визначаються параболоїди.
Введемо в розгляд символи  та
та  , означивши їх рівностями
, означивши їх рівностями
 ,
,
 ,
,
 ,
,
а також
 ,
,  ,
,  .
.
Вираз  є половиною похідної від функції
є половиною похідної від функції  по змінній
по змінній  , якщо змінні
, якщо змінні  та
та  при цьому вважати сталими. Аналогічно,
при цьому вважати сталими. Аналогічно,  та
та  - це половини похідних від функції по змінних та відповідно при умові, що дві інші змінні вважаються сталими.
- це половини похідних від функції по змінних та відповідно при умові, що дві інші змінні вважаються сталими.
Найближчими нашими задачами буде дослідження властивостей поверхонь, заданих загальним рівнянням, вивчення особливостей їх розташування відносно системи координат, а також дослідження питання, скільки та які різні види поверхонь може визначати рівняння (1).
2. Перетнемо поверхню другого порядку прямою  , яка проходить через деяку точку
, яка проходить через деяку точку  паралельно до вектора
паралельно до вектора  . Запишемо параметричні рівняння прямої у виді
. Запишемо параметричні рівняння прямої у виді
 (2)
(2)
та знайдемо точки перетину поверхні та прямої . Дістаємо систему рівнянь (1), (2), розв’язуючи яку відносно змінної  , отримуємо квадратне рівняння
, отримуємо квадратне рівняння
 , (3)
, (3)
де
 ,
,





 ,
,  Дослідимо особливості взаємного розташування поверхні та прямої у випадках, коли деякі з коефіцієнтів рівняння (3) перетворюються в нуль.
Дослідимо особливості взаємного розташування поверхні та прямої у випадках, коли деякі з коефіцієнтів рівняння (3) перетворюються в нуль.
1. Нехай  . Одному із коренів рівняння
. Одному із коренів рівняння  , який рівний нулю, відповідає точка . Тому у цьому випадку одна із точок перетину поверхні та прямої співпадає з точкою
, який рівний нулю, відповідає точка . Тому у цьому випадку одна із точок перетину поверхні та прямої співпадає з точкою  (рис. 1).
(рис. 1).
 2. Нехай
2. Нехай  та рівняння
та рівняння  має два дійсні корені
має два дійсні корені  . Цим кореням відповідають дві точки, які належать заданій поверхні та прямій – це точки
. Цим кореням відповідають дві точки, які належать заданій поверхні та прямій – це точки  . У цьому випадку точка є серединою хорди
. У цьому випадку точка є серединою хорди  (рис. 2). Якщо дане рівняння має два уявні корені, то пряма буде перетинати поверхню у двох уявних точках, а точка буде серединою уявної хорди.
(рис. 2). Якщо дане рівняння має два уявні корені, то пряма буде перетинати поверхню у двох уявних точках, а точка буде серединою уявної хорди.
3. Нехай  . Рівняння
. Рівняння  має єдиний корінь
має єдиний корінь  , який визначає першу із точок перетину. Щоб зрозуміти особливість розташування другої точки перетину доцільно дослідити, як змінюється другий корінь рівняння (3) при
, який визначає першу із точок перетину. Щоб зрозуміти особливість розташування другої точки перетину доцільно дослідити, як змінюється другий корінь рівняння (3) при  . У лекції 18, де досліджувались аналогічні питання взаємного розташування лінії другого порядку та прямої, було показано, що при абсолютна величина другого кореня прямує до
. У лекції 18, де досліджувались аналогічні питання взаємного розташування лінії другого порядку та прямої, було показано, що при абсолютна величина другого кореня прямує до  . Згідно із рівностями (2) при
. Згідно із рівностями (2) при  друга із точок перетину нескінченно віддаляється від точки . Таку точку ми, аналогічно до попереднього, будемо позначати символом
друга із точок перетину нескінченно віддаляється від точки . Таку точку ми, аналогічно до попереднього, будемо позначати символом  та говорити, що пряма перетинає поверхню у нескінченно віддаленій точці. Напрям прямої при цьому будемо називати асимптотичним. Асимптотичним буде, наприклад, напрям прямої, яка перетинає еліптичний параболоїд та проведена паралельно до його осі симетрії (рис.
та говорити, що пряма перетинає поверхню у нескінченно віддаленій точці. Напрям прямої при цьому будемо називати асимптотичним. Асимптотичним буде, наприклад, напрям прямої, яка перетинає еліптичний параболоїд та проведена паралельно до його осі симетрії (рис.  ), або прямої, яка проведена паралельно до твірної конуса (рис.
), або прямої, яка проведена паралельно до твірної конуса (рис.  ).
).

 4. Випадок
4. Випадок  є поєднанням розглянутих вище випадків 1, 2. Рівняння (3) матиме вид
є поєднанням розглянутих вище випадків 1, 2. Рівняння (3) матиме вид  та корені
та корені  . У цьому випадку пряма буде дотикатись до поверхні у точці (рис. 4).
. У цьому випадку пряма буде дотикатись до поверхні у точці (рис. 4).
 5. При
5. При  точка належить поверхні , а пряма матиме відносно асимптотичний напрям.
точка належить поверхні , а пряма матиме відносно асимптотичний напрям.
6. Якщо  , то рівняння (3) не має розв’язків. У цьому випадку пряма не має з поверхнею спільних точок та має відносно асимптотичний напрям.
, то рівняння (3) не має розв’язків. У цьому випадку пряма не має з поверхнею спільних точок та має відносно асимптотичний напрям.
7. Якщо  , то розв’язком рівняння (3) буде довільне дійсне число . Тоді кожна точка прямої належить поверхні . Ми зустрічалися із таким випадком, коли говорили про прямолінійні твірні поверхонь другого порядку, всі точки яких належать поверхні.
, то розв’язком рівняння (3) буде довільне дійсне число . Тоді кожна точка прямої належить поверхні . Ми зустрічалися із таким випадком, коли говорили про прямолінійні твірні поверхонь другого порядку, всі точки яких належать поверхні.
3.Хордою поверхні другого порядку назвемо відрізок, який сполучає дві її довільні точки.
Якщо існує точка, в якій усі хорди, які проходять через неї, діляться пополам, то цю точку називають центром поверхні.
Центр поверхні є її центром симетрії, оскільки разом із будь-якою точкою поверхні їй належить також точка, симетрична даній відносно центра.
Розглянемо питання відшукання центра поверхні, заданої рівнянням (1). Як ми знаємо, умовою того, щоб точка  була серединою хорд, які проходить через неї, згідно пунктом 2), є виконання рівності
була серединою хорд, які проходить через неї, згідно пунктом 2), є виконання рівності
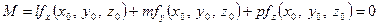
для довільного напрямку, який задається вектором . Тому для відшукання центра лінії дістаємо систему рівнянь
 . (4)
. (4)
Існування та кількість розв’язків системи (4) залежить від її визначника  .
.
Якщо  , то система (4) має єдиний розв’язок. У цьому випадку поверхня має єдиний центр і її називають центральною. Прикладами таких поверхонь є еліпсоїд, одно та двопорожнинний гіперболоїд, конус.
, то система (4) має єдиний розв’язок. У цьому випадку поверхня має єдиний центр і її називають центральною. Прикладами таких поверхонь є еліпсоїд, одно та двопорожнинний гіперболоїд, конус.
Якщо  , то система (4) має безліч, або не має жодного розв’язку. Поверхню у цьому випадку називають нецентральною. Прикладами поверхонь, які не мають жодного центра, є еліптичний та гіперболічний параболоїди, параболічний циліндр. Якщо система (4) має безліч розв’язків, то в залежності від рангу її матриці відповідні розвя’зкам точки утворюють пряму або площину. Еліптичний та гіперболічний циліндри мають пряму центрів, а поверхня, яка вироджується у дві паралельні площини – площину центрів. Ця площина проходить паралельно до даних двох площин та знаходиться від них на однаковій відстані.
, то система (4) має безліч, або не має жодного розв’язку. Поверхню у цьому випадку називають нецентральною. Прикладами поверхонь, які не мають жодного центра, є еліптичний та гіперболічний параболоїди, параболічний циліндр. Якщо система (4) має безліч розв’язків, то в залежності від рангу її матриці відповідні розвя’зкам точки утворюють пряму або площину. Еліптичний та гіперболічний циліндри мають пряму центрів, а поверхня, яка вироджується у дві паралельні площини – площину центрів. Ця площина проходить паралельно до даних двох площин та знаходиться від них на однаковій відстані.
Приклад 1. Знайти центри поверхні, заданої рівнянням  .
.
Розв’язання. Складемо та розв’яжемо систему рівнянь  . Дістаємо
. Дістаємо  , звідки
, звідки  . Отже, задана поверхня має єдиний центр, який знаходиться у точці
. Отже, задана поверхня має єдиний центр, який знаходиться у точці  .
.
4. Згідно з попереднім, пряма буде дотичною до поверхні , якщо для рівняння , яке характеризує перетин прямої з поверхнею , виконуються умови  . Нехай точка належить поверхні, тобто
. Нехай точка належить поверхні, тобто  . Всі прямі, які проходять через точку
. Всі прямі, які проходять через точку  та дотикаються до поверхні, утворюють площину, оскільки незалежно від напрямку вектора вони перпендикулярні до сталого вектора
та дотикаються до поверхні, утворюють площину, оскільки незалежно від напрямку вектора вони перпендикулярні до сталого вектора  . Справді, з рівності , яку можна записати у виді
. Справді, з рівності , яку можна записати у виді
 ,
,
випливає, що 

 . Цю площину називають дотичною площиною до поверхні. Знайдемо рівняння такої площини. Оскільки відомий вектор , який перпендикулярний до площини, а також точка , яка їй належить, то рівняння дотичної площини запишеться у виді
. Цю площину називають дотичною площиною до поверхні. Знайдемо рівняння такої площини. Оскільки відомий вектор , який перпендикулярний до площини, а також точка , яка їй належить, то рівняння дотичної площини запишеться у виді
 . (5)
. (5)
Нормаллю до поверхні, проведеній в деякій її точці, називають пряму, яка перпендикулярна до дотичної площини у цій же точці. Оскільки вектор паралельний до нормалі, яка проведена у точці , то рівняння останньої запишеться у виді
 . (6)
. (6)
Приклад 2. На поверхні  знайти точки, в яких нормаль буде паралельною до осі
знайти точки, в яких нормаль буде паралельною до осі  .
.
Розв’язання. Оскільки вектор паралельний до осі , то  . Обчислюємо
. Обчислюємо 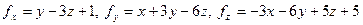 .
.
Розв’язуючи систему  знаходимо шукані точки:
знаходимо шукані точки:  .
.
Відповідь: .
Л Е К Ц І Ї
З
АНАТОМІЇ ТА ЕВОЛЮЦІЇ
НЕРВОВОЇ СИСТЕМИ
Миколаїв – 2010
Вчення про нервову систему (неврологія)
Однією із властивостей живого є подразливість. Кожний живий організм отримує подразнення від навколишнього середовища і відповідає на них відповідними реакціями, які пов’язують організм із зовнішнім світом. Обмін речовин, який відбувається в самому організмі, у свою чергу, зумовлює низку подразнень, на які реагує організм. Зв’язок між ділянкою, на яку падає подразнення, і реагуючим органом у вищому багатоклітинному організмі здійснюється нервовою системою.
Вона проникає у всі відділи тіла людини та інтегрує органи в їх системи, а системи органів – у цілісний організм, і відповідає за взаємодію організму людини з навколишнім світом. Отже, нервова система виконує наступні функції: 1) об’єднуючу (інтегративну); 2) регулюючу – регулює функції органів; 3) зумовлює взаємозв’язок між організмом і навколишнім середовищем.
Єдина нервова система людини поділяється на дві основних складових частини: 1) соматичну та 2) вегетативну. Соматична частина нервової системи керує посмугованою м’язовою тканиною скелета і деяких внутрішніх органів (язика, глотки, гортані) та містить у своєму складі центри і провідники чутливості всього тіла. Вегетативна частина забезпечує рухову і секреторну іннервацію всіх внутрішніх органів (травна, дихальна, сечостатева системи) та шкіри, а також іннервує серце і судини. У свою чергу вегетативну частину нервової системи поділяють на симпатичний і парасимпатичний відділи.
У соматичній і вегетативній частинах нервової системи розрізняють центральний відділ – головний і спинний мозок та периферичний – черепно-мозкові і спинномозкові нерви (з вузлами на їх задніх корінцях), пограничний симпатичний стовбур, його гілки і численні вегетативні вузли у стінках внутрішніх органів і біля них.
Умовність і обмеженість наведеної класифікації очевидна. Вегетативна нервова система має відношення до органів соми, наприклад, до скелетних м’язів, а у центральному відділі соматичної нервової системи розміщені центри її вегетативної частини. Всі функції вегетативної і соматичної нервової системи підпорядковані корі головного мозку і нею регулюються.
Головний мозок, його кора і тісно пов’язані з нею підкіркові утворення є матеріальною основою вищої нервової діяльності.
Структурно-функціональною одиницею, або найдрібнішою часточкою органу, здатною виконувати його функцію, в нервовій системі є нейрон (нейроцит, нервова клітина).
Нервова клітина складається з тіла, яке містить ядро із цитоплазмою і органелами, та з відростків. Розрізняють короткі відростки, які розгалужуються, – дендрити і довгий відросток – аксон (осьовий циліндр, нейрит). Нейрон функціонує у тісному зв’язку з оточуючою його глією і кровоносними судинами. Уся нервова система складається із ланцюгів нейронів і гліальної строми; нейрони з’єднуються між собою за допомогою контактів – синапсів.
Синапс — спеціалізований контакт між нервовими клітинами або між нервовою клітиною та м’язом, у зоні якого відбувається передача нервового імпульсу. Основні структурні компоненти синапсу: пресинаптична мембрана (ділянка плазмолеми відростка нервової клітини, з якої надходить сигнал), постсинаптична мембрана (ділянка плазмолеми клітини, яка сприймає сигнал), синаптична щілина шириною 20-30 нм (розмежовує пре- і постсинаптичну мембрани), заповнені нейромедіатором синаптичні пухирці. Медіатором називається біологічно активна речовина, нейромедіатором – та, яка викликає відповідну реакцію нервової системи. Функціонування синапсів забезпечує односторонню передачу інформації від клітини до клітини за допомогою медіатора (хімічного посередника). За хімічною будовою медіаторами є ацетилхолін, адреналін, норадреналін та інші. Виходячи із цього, синапси бувають адренергічними та холінергічними.
Розрізняють три сторони діяльності нервової системи: 1) рецепторну функцію – сприйняття подразнення і розповсюдження нервового імпульсу по нервових провідниках до центру, з цього явища починається аналіз; 2) замикаючу функцію – процес перетворення отриманого нервовим центром імпульсу в певні зовнішні реакції, з нього починається синтез; 3) ефекторну, рухову функцію – здійснення відповідної реакції (рухової або секреторної).
Відповідно до цього є три основні типи нейронів: 1) рецепторні (аферентні, чутливі) нейрони. Розрізняють наступні типи рецепторів: а) екстероцептори, які сприймають подразнення із зовнішнього середовища; б) інтерцептори, які сприймають подразнення від внутрішніх органів і судин (включаючи і больову чутливість); в) пропріоцептори – один із різновидів інтерцепторів, сприймає подразнення в органах опори і руху (в м’язах, суглобах, сухожилках, окісті – “м’язово-суглобове відчуття”). Їх тіла завжди лежать поза межами центральної нервової системи. Периферична частина несправжньооднополюсного нейрону (а саме із таких нейронів побудовані рецепторні нейрони) направляється на периферію, закінчуючись там певним чутливим закінченням; центральна частина направляється у спинний мозок чи у стовбурову частину головного мозку; 2) замикаючі (асоціативні) нейрони, які лежать у межах центральної нервової системи; 3) ефекторні (еферентні, рухові) нейрони. Їх тіла знаходяться у центральній нервовій системі (або у вегетативних вузлах), а нейрити (аксони), продовжуючись у вигляді осьових циліндрів нервових волокон, досягають робочих органів (м’язи, залози).
Поряд із функціональною класифікацією нейронів існує і морфологічна:
1) однополюсні (уніполярні) нейрони, 2) двополюсні (біполярні), 3) багатополюсні (мультиполярні) нейрони, 4) несправжньооднополюсні (псевдоуніполярні) нейрони.
Філогенез нервової системи
Гуморальна, донервова форма регуляції присутня у всіх одноклітинних організмів. Так, у найпростіших одноклітинних організмів (амеби) нервової системи немає. Регуляція відправлень клітини здійснюється гуморальним шляхом – біологічно активні речовини через цитоплазму клітини поступають до певної ділянки клітини і здійснюють свій регулюючий вплив.
Нервова форма регуляції виникла з появою багатоклітинних організмів, у яких з’явилася нервова система. У своєму розвитку нервова система проходить три етапи.
І етап – сіткоподібна нервова система, приклад – кишковопорожнинні (гідра). Її нервова система складається з нервових клітин, численні відростки яких, багаторазово з’єднуючись між собою у різних напрямках, утворюють сітку, яка дифузно пронизує все тіло. При подразненні будь-якої ділянки тіла збудження розповсюджується по всій нервовій сітці, і тварина реагує рухом усього тіла. Відображенням цього етапу в людини є сіткоподібна будова інтрамуральної нервової системи.
ІІ етап – вузлова нервова система. Нервові клітини уже утворюють вузли і нервові стовбури. Поперечні нервові стовбури з’єднують вузли в межах сегменту, поздовжні нервові стовбури різних сегментів зв’язують нервові вузли сусідніх сегментів у єдине ціле. На головному кінці, який при рухові дотикається до різних предметів оточуючого середовища, розвиваються органи чуттів, у зв’язку з чим головні вузли розвиваються сильніше за решту, виступаючи прообразом майбутнього головного мозку. Відображенням цього етапу є збереження у людини примітивних рис (розкиданість по всій території вузлів і мікровузлів) у будові вегетативної нервової системи.
ІІІ етап – трубчаста нервова система. На початковому ступені розвитку тварин особливо велику роль відігравав апарат руху, від досконалості якого залежала основна умова існування – живлення (рух у пошуках їжі, захват її та поглинання).
У нижчих хребетних розвився перистальтичний спосіб переміщення, пов’язаний з непосмугованими м’язами і місцевим нервовим апаратом. На більш високому ступені розвитку перистальтичний спосіб змінюється скелетною моторикою – рухом за допомогою жорстких важелів – поверх м’язів (членистоногі) і всередині м’язів (хребетні). Наслідком цього стало утворення посмугованої м'язової тканини і центральної нервової системи, яка координує переміщення окремих важелів моторного скелета. Так нервова система у хордових (ланцетник) виникла у вигляді метамерно побудованої нервової трубки із сегментарними нервами, які від неї відходять до всіх сегментів тіла, включно з апаратом руху – тулубний мозок. У хребетних і людини тулубний мозок стає спинним. Таким чином, поява тулубного мозку пов’язана з удосконаленням у першу чергу моторного апарату тварини. Поряд із цим уже в ланцетника є і рецептори (нюховий, світловий).
Подальший розвиток нервової системи і виникнення головного мозку зумовлені переважно вдосконаленням рецепторного апарату.
Оскільки більшість органів чуттів виникає на тому кінці тіла, який звернений у бік руху (вперед), то для сприйняття зовнішніх подразнень, які до них поступають, розвивається передній кінець тулубного мозку і утворюється головний мозок, що співпадає з відокремленням переднього кінця тіла у вигляді голови – цефалізація (cephalic – голова).
На першому етапі розвитку головний мозок складається із трьох відділів: переднього, середнього та заднього, причому із цих відділів у першу чергу (в нижчих риб) особливо розвивається задній, або ромбоподібний мозок. Розвиток заднього мозку відбувається під впливом рецепторів акустики і статики (рецептори VIII пари черепних нервів), які мають вирішальне значення для орієнтації у водному середовищі.
У подальшій еволюції задній мозок диференціюється на довгастий мозок, який є перехідним відділом від спинного мозку до головного, та власне задній мозок, із якого розвиваються мозочок і міст.
У процесі пристосування організму до навколишнього середовища шляхом зміни обміну речовин у задньому мозку як найбільш розвиненому відділі центральної нервової системи виникають центри керування життєво важливими процесами рослинного життя, пов’язані, зокрема, із зябровим апаратом (дихання, кровообіг, живлення). Тому в довгастому мозку з’являються ядра зябрових нервів (Х пара – блукаючий нерв). Ці життєво важливі центри дихання і кровообігу залишаються у довгастому мозку людини, чим пояснюється смерть при ушкодженні довгастого мозку.
На третьому етапі, у зв’язку з остаточним переходом тварин із водного середовища в повітряне, посилено розвивається нюховий рецептор, який сприймає хімічні речовини, що містяться у повітрі. Вони сигналізують своїм запахом про здобич, небезпеку та інші життєво важливі явища навколишнього середовища. Під впливом нюхового рецептора розвивається передній мозок. У подальшому передній мозок розростається і диференціюється на проміжний та кінцевий мозок. У кінцевому мозку як у вищому відділі центральної нервової системи з’являються центри усіх видів чутливості. Однак нижче розміщені не зникають, а зберігаються, підпорядковуючись вище розміщеним центрам. Отже, з кожним новим розвитком головного мозку виникають нові центри, які підкорюють собі старі.
Відбувається своєрідне переміщення функціональних центрів до головного кінця і одночасне підпорядкування філогенетично старих зачатків новими. У результаті центри слуху, які вперше виникли у задньому мозку, є також у середньому і передньому. Центри зору, які виникли у середньому мозку, є також і в передньому, а центри нюху – тільки в передньому мозку.
Вдосконалення рецепторів призводить до прогресивного розвитку переднього мозку, який поступово стає органом, що керує всією поведінкою тварини.
Розрізняють дві форми поведінки тварин: інстинктивну, яка основана на видових реакціях (безумовні рефлекси), та індивідуальну, основану на досвіді індивідуума (умовні рефлекси). Відповідно до цих двох форм поведінки у кінцевому мозку є дві групи центрів сірої речовини: підкіркові вузли, які мають форму ядер (ядерні центри), та кора з сірої речовини, яка має будову суцільного екрану (екранні центри). При цьому спочатку розвиваються підкіркові ядра, а потім кора. Кора виникає при переході тварин від водного до наземного способу існування і чітко виявляється в амфібій і рептилій.
Подальша еволюція нервової системи характеризується тим, що кора головного мозку все більше підпорядковує собі нижчерозміщені центри, відбувається поступова кортиколізація функцій.
Необхідною формацією для здійснення вищої нервової діяльності є нова кора, розміщена на поверхні півкуль, яка набула у процесі філогенезу шестишарової будови. Завдяки посиленому розвитку нової кори кінцевий мозок у вищих хребетних переважає решту відділів головного мозку, нібито покриваючи їх плащем (pallium). Новий мозок (neencephalon), який розвився, відтісняє у глибину старий (нюховий) мозок, який згортається у вигляді Аммонового рогу (hyppocamp), який залишається нюховим центром. Внаслідок плащ (новий мозок) різко переважає над рештою відділів головного мозку – старим мозком (pallencephalon).
Отже, розвиток головного мозку здійснюється під впливом розвитку рецепторів, чим і пояснюється, що найвищий відділ головного мозку – кора сірої речовини, являє собою сукупність кіркових кінців аналізаторів, суцільну рецепторну поверхню, яка сприймає подразнення. Подальший розвиток мозку в людини піддається іншим закономірностям, пов’язаним із його соціальною природою.
Друга сигнальна система
Крім аналізаторів, які сприймають різноманітні подразнення зовнішнього світу і складають конкретно – наочне мислення, властиве тваринам (перша сигнальна система), у людини виникла здатність абстрактного мислення, за допомогою слова, спочатку почутого (усна мова), пізніше видимого (письмова мова). Це склало другу сигнальну систему.
Кіркові центри аналізаторів мови:
1. Руховий аналізатор артикуляції мови, центр Брока, знаходиться у задній частині верхньої скроневої звивини. При його враженні спостерігається рухова афазія – людина не може вимовити слова.
2. Слуховий аналізатор усної мови. Його ядро розміщене в задній частині верхньої скроневої звивини, в глибині бічної (Сильвієвої) борозни – центр Верніке. При його враженні зберігається здатність чути звуки, але втрачається властивість розуміти слова – словесна глухота або сенсорна афазія.
3. Руховий аналізатор писемної мови розміщується у задньому відділі середньої лобної звивини. При враженні цього центру втрачається здатність писати – аграфія.
4. Зоровий аналізатор письмової мови розміщений у нижній тім’яній часточці в ділянці кутової звивини. При його враженні людина втрачає здатність читати – алексія.
Всі аналізатори мови закладаються у обох півкулях, але розвиваються тільки з одного боку (у правші – зліва, у лівші – справа).
Аналізатори усної і писемної мови сприймають словесні сигнали, що складає другу сигнальну систему дійсності у формі абстрактного мислення (загальні уявлення, поняття, висновки, узагальнення), властиві тільки людині.
Відомо, що тіло людини побудоване за типом двохсторонньої (білатеральної) симетрії і що воно, в основному, поділяється на сегменти (метамери). Ці особливості будови добре виражені і в ділянці нервової системи, перш за все у спинному мозкові. За зовнішніми ознаками спинний мозок людини не виявляє ніяких ознак сегментації: це довгий, приблизно циліндричної форми тяж, але всередині він містить сегментарні центри, з якими пов’язані у метамерному порядку спинномозкові нерви. У головному мозкові розрізняють два основних відділи: 1) мозковий стовбур, який зберігає сліди метамерії і є безпосереднім видозміненим спинним мозком (до мозкового стовбура відносяться довгастий мозок, міст, мозочок, середній мозок, проміжний мозок) 2) півкулі великого мозку.
Короткий порівняноанатомічний нарис спинного мозку
Спинний мозок зазвичай має форму довгого тяжа, розміщеного у хребтовому каналі на протязі шиї, всього тулуба і хвостової ділянки. У нижчих хребетних спинний мозок за довжиною дорівнює довжині хребтового канала, але вже у деяких Teleostei він укорочуються. У Anura i Mammalia його каудальна частина являє собою тонку кінцеву нитку, filum terminale. В задньому відділі проходять тільки корінці поперекових і крижових нервів, які утворюють так званий кінський хвіст. Особливо значна невідповідність між довжиною спинного мозку і хребтового канала у ссавців.
Спинний мозок у багатьох тварин утворює два потовщення – шийне і поперекове, які розміщені там, де відходять найтовстіші нерви (до кінцівок). Ці потовщення, intumescentiae, краще за все виражені в Anura, Chelonia, Aves і Mammalia. Спинний мозок риб, безногих амфібій, змій і безногих ящірок не має потовщень.
Внутрішня будова спинного мозку. У Cyclostomata біла речовина складається тільки із безмієлінових волокон. Нервові клітини починають поступово концентруватись біля центрального канала, утворюючи сіру речовину, але у риб вона не відмежована від білої; навіть у амфібій дорзальні роги спинного мозку розвинені слабко; в той час, як вентральні уже виражені добре. У Sauropsida і Mammalia роги сірої речовини добре розвинені, у зв’язку з чим біла речовина поділяється на поздовжні передні, задні та бічні канатики, funicli anteriores, рosteriorеs et laterales. У міру накопичення нервових волокон у спинному мозку не тільки потовщуються стінки центрального канала, але все більш звужується його просвіт, особливо у дорзальній частині; тут стінки канала поступово наближаються одна до одної і зливаються. На цьому місці розвивається серединна пластинка гліальної тканини – спинна перегородка. Бічні стінки потовщуються значно сильніше, ніж дорзальна і вентральна. Метамерія у спинному мозку виражається у групуванні по сегментам нервових клітин у ділянці вентральних і дорзальних рогів сірої речовини, а зовні – відходженням через певні проміжки корінців спинномозкових нервів.
Особливість спинного мозку ссавців і людини – у волокнах пірамідних шляхів – передніх і бічних, які безпосередньо зв’язують кору півкуль з клітинами передніх рогів. Маса цих волокон зростає від нижчих до вищих, у зв’язку з поступовим збільшенням (у філогенезі) плащового відділу головного мозку: так, у собаки їх кількість складає приблизно 10% всієї білої речовини спинного мозку, у мавп – 20%, у людини – майже 30%. Це свідчить про все збільшуючий вплив кори півкуль на роботу м’язів, що збільшуються.
Короткий нарис ембріогенезу спинного мозку
Закладка нервової системи проходить із ектодерми у вигляді її потовщення – нервової пластинки, яка потім перетворюється на нервову (мозкову) трубку. Клітини останньої диференціюються у двох напрямках: одні – спонгіобласти, диференціюються в елементи нейроглії; інші– нейробласти – перетворюються на нейрони.
Із краніального відділу нервової трубки утворюється головний мозок, із каудального її відділу – спинний мозок.
Спочатку всі стінки каудального відділу мозкової трубки мають однакову товщину. В подальшому її бічні стінки все більше потовщуються, тоді як вентральна і дорзальна, відстаючи у рості, поступово йдуть у глибину, а на їх місці по серединній площині утворюються вентральна і дорзальна поздовжні борозни, які поділяють спинний мозок на дві симетричні половини.
На внутрішній поверхні кожної бічної стінки також намічається по одній поздовжній (пограничній) борозенці, яка поділяє ці стінки на два відділи: вентральний (основна пластинки) і дорзальний (крилова пластинка).
З основної пластинки утворюються передні стовпи сірої речовини спинного мозку з білою речовиною, яка прилягає до нього. Із крилової пластинки розвиваються задні стовпи сірої речовини і суміжна із ним біла речовина. Нейрити (аксони) нервових клітин, які утворюються в основних пластинках, покидаючи спинний мозок, складають його передні корінці. Нейрити клітин спінальних гангліїв, які вступають у ділянці задніх стовпів спинного мозку, утворюють його задні корінці.
При замиканні нервової пластинки у трубку в її відділах, які переходять безпосередньо в ектодерму, утворюється гангліозна пластинка, яка йде у вигляді тяжа по дорзальній стінці мозкової трубки. Пізніше це утворення розщеплюється на всю довжину, обидві його половини переміщуються вентрально, розміщуючись вздовж із боків мозкової трубки у вигляді нервових валиків. Обидва валики в подальшому багаторазово перешнуровуються, відповідно до сегментів дорзальної сторони зародка; у результаті чого на кожному боці утворюється низка міжхребцевих вузлів. Із останніх виділяються вертебральні та інші вузли симпатичної нервової системи.
Короткий порівняноанатомічний нарис головного мозку
Майже у всіх Craniota на ранніх стадіях зародкового періоду головний кінець мозкової трубки диференціюється на три первинних мозкових пухирі: передній – prosencephalon, середній – mesencephalon, ромбоподібний – rhombencephalon. У подальшому передній і ромбоподібний пухирі знову поділяються, кожний на дві частини. Отже, утворюється п’ять відділів (починаючи спереду): кінцевий мозок – telencephalon, проміжний мозок – diencephalon, середній мозок – mesencephalon, задній мозок – metencephalon, довгастий мозок – myelencephalon; останні два – із ромбоподібного мозку. Центральний канал спинного мозку, переходячи у головний, розширюється у систему шлуночків мозку, які з’єднуються між собою. Розподіл сірої та білої речовини у головному мозкові у нижчих черепних в основному такий же, як і у спинному: шлуночки вистелені сірою речовиною, біла лежить зовні; та й інша є продовженням відповідних шарів спинного мозку. Крім того, у мозочку, а у вищих і в кінцевому мозку (півкулі великого мозку) зовні від білої речовини з’являється і сіра – кора мозку, cortex cerebri.
Cyclostomata. Головний мозок примітивний. Довгастий мозок, дуже схожий за будовою на спинний мозок, але в ньому вже є ромбоподібна ямка. Мозочок і середній мозок розвинені слабко; в останньому намічається поділ на два горби. Проміжний мозок побудований складніше: бічні стінки третього шлуночка потовщуються у зорові горби, нижня стінка тонка, тут утворюється вентральне випинання – лійка, дах має два непарних випинання, за будовою – рудиментарні органи зору: заднє більше – пінеальний орган, переднє – тім’яний, або парапінеальний орган. Кінцевий мозок розвинений слабко, складається із двох симетричних половинок, кожна поділяється на два відділи: задній – власне півкуля, передній більший – нюхова доля, порожнина її з’єднуються із бічними шлуночками.
Головну масу півкуль складають сильно розвинені смугасті тіла. Верхня стінка півкуль тонка, складається із низки епітеліальних клітин, у міксин зовсім відсутні нервові елементи; у міног містить на обмеженому протязі пірамідні клітини – перший зачаток плащу.
В селахій довгастий мозок значної довжини, має низку поздовжніх підвищень, які містять ядра черепномозкових нервів. Мозочок інколи за розміром дорівнює передньому мозку, часто поділяються поперечними борозенками на часточки. Середній мозок також розвинений добре, поділяється серединною борозною на два горби. У проміжному мозку з дорзального боку розвинений тільки епіфіз. Кінцевий мозок іще не поділений на півкулі, містить непарний шлуночок, спереду переходить у парні нюхові долі. Стінка кінцевого мозку складається з епітелію та нервової тканини. У ганоїд і костистих дуже розвинений середній мозок; його дорзальна частина – покришка зазвичай складається із двох горбів. Головна маса кінцевого мозку представлена смугастими тілами, поділеними на дорзальну частину – верхнє смугасте тіло, епістріатум, та вентральну – власне смугасте тіло, corpus striatum. Плащовий відділ – у вигляді тонкої епітеліальної пластинки, інколи містить невелику кількість нервової тканини (ганоїди). Поділ кінцевого мозку на півкулі слабо помітний, порожнина шлуночка спільна. Кінцевий мозок продовжується безпосередньо у нюхові часточки.
У Dipnoi кінцевий мозок переважає решту відділів, складається із двох півкуль, стінки їх містять нервову тканину, яка вистеляє порожнини шлуночків; тут можна вже говорити про розвиток древньої кори – archipallium. Середній мозок (непарний горб) значно поступається передньому мозку. Мозочок дуже малий.
Amphibia. Довгастий мозок скорочений. Мозочок у вигляді вузької пластинки. Середній мозок складається із двох горбів. Є шишкоподібне тіло, тім’яний орган відсутній. Зорові горби добре виражені. У підталамічній ділянці є об’ємний гіпофіз. Кінцевий мозок переважає решту відділів, складається із двох подовжених півкуль, які переходять спереду у нюхові частки. У півкулях уже добре виражений плащовий відділ. Частина нервових клітин переміщується із вистеляючої порожнини шлуночків центральної сірої речовини до периферії і утворює зачаток кори – archicortex, у вигляді окремих гнізд. Смугасті тіла виражені порівняно слабко.
Рептилії. Мозочок розвинений сильніше, ніж у амфібій, але значної величини досягає тільки у крокодилів; у ньому, окрім непарної середньої частини, намічаються і бічні – flocculi (майбутні півкулі мозочка). Середній мозок представлений у вигляді двох горбів, до яких у крокодилів приєднується два задніх. Проміжний мозок із дорзального боку покритий сусідніми відділами (кінцевим і середнім мозком). Зорові горби відносно великі, є шишкоподібне тіло. У деяких рептилій (Sauria, Hatteria) добре виражений тім’яний орган – рудиментарний орган зору. Кінцевий мозок складається із нюхових часток і добре розвинених півкуль; останні складаються з ясно виражених смугастих тіл і плащового відділу. У плащовому відділі добре розвинений поверхневий шар сірої речовини із пірамідними клітинами, це – вже істинна кора великого мозку.
Aves. Спільна маса головного мозку значно збільшується. Довгастий мозок коротший, ніж у рептилій. Мозочок представлений черв’яком, досягає дуже великих розмірів, його поверхня зростає завдяки появі поперечних борозен, півкулі його знаходяться у зачатковому стані. Середній мозок складається із двох великих зорових часток. Проміжний мозок схований за півкулями і мозочком; між ними видно лише епіфіз, який добре розвинений. Кінцевий мозок у птахів – відносно більших розмірів, головну його масу складають смугасті тіла дуже складної будови. Намічаються скронева, лобна і потилична частки. Кора розвинена слабко. Структура кори стає більш складною. Нюхові частки незначні.
Mammalia. Головний мозок ссавців характеризується особливо високим розвитком плащового відділу і диференціацією кори. Довгастий мозок короткий, у його вентральній частині розвиваються піраміди, латеральніше від яких розміщені у вигляді підвищень нижні оливи; вони вже є у риб, але у вищих ссавців випинаються на поверхню. Пучки волокон, які відмежовують каудальний відділ ромбоподібної ямки, продовжуються у вигляді вірьовчаних тіл у мозочок, утворюючи його задні ніжки. На вентральній поверхні переднього відділу довгастого мозку у ссавців з'являється нова спайка – міст, pons, пучки якого ідуть до півкуль мозочка, утворюючи його середні ніжки. Мозочок розвинений добре, складається із середньої непарної частини – черв’яка та бічних – півкуль. Ці утворення, особливо у вищих ссавців, поділені на велику кількість часточок, за рахунок яких поверхня мозочка збільшується.
Середній мозок, окрім верхніх двох горбків, типових для нижчих хребетних, містить іще два нижніх горбка, colliculus inferior – утворюється покрівля середнього мозку. Із зоровим нервом пов’язані тільки верхні горбки, нижні – зі слуховим нервом. Стінки середнього мозку потовщуються, його порожнина перетворюється на вузький канал – водопровід середнього мозку, aqueductus mesencephali. Вентральна частина середнього мозку утворена ніжками мозку, які містять пучки нервових волокон і скупчення нервових клітин (ядра).
Проміжний мозок розвинений відносно добре; основну його масу утворюють зорові горби (згір’я), в них у нижчих ссавців закінчується більша частина зорових волокон. Тім’яний орган відсутній, епіфіз майже завжди є. У підзгірній ділянці, назад від перехрестя, розміщується лійка із гіпофізом на кінці; задня стінка лійки має ознаки сірого горба. У його основи знаходиться непарне підвищення – сосочкове тіло, у приматів – парне. Назад від згір’я, на межі із середнім мозком – два парних підвищення – колінчаті тіла; у приматів бічні колінчаті тіла приймають велику кількість зорових волокон. Медіальні колінчаті тіла намічаються ще у птахів, але зовні їх не видно; вони приймають волокна від слухового нерва.
Кінцевий мозок –найхарактерніша частина мозку ссавців, найбільш сильний ріст виявляє плащовий відділ (pallium), тоді як вузли основи збільшуються порівняно мало; вони тут поділяються пучками нервових волокон на окремі ядра; з них найголовніші – хвостате і сочевицеподібне. Мигдалеподібне , гомологічне епістріатуму нижчих, зміщується у ділянку нижнього рогу.
Півкулі розростаються у всіх напрямках, утворюються окремі частки: лобна, тім’яна, скронева, потилична; перші дві більш раннього походження; порожнини бічних шлуночків простягаються у всі частки у вигляді рогів. Від нижчих до вищих ріст півкуль прогресує; вони поступово покривають все більшу частину мозкового стовбура. При розгляді головного мозку з дорзального боку проміжний мозок схований у всіх ссавців. Середній мозок видимий зовні у багатьох нижчих плацентарних. У копитних, хижаків, китоподібних і приматів він уже покритий заднім відділом півкуль, а у людиноподібних і у людей останній схований зовсім.
Початково (наприклад, у круглоротих) нюхові частки складають головну масу кінцевого мозку; філогенетично зачаток кінцевого мозку був пов’язаний саме із органом нюху. Серед ссавців нижчі мають добре розвинені нюхові частки, які виступають спереду від півкуль. У вищих приматів нюховий мозок (rhinencephalon), редукується; нюхові частки мають незначні придатки, покриті лобними частками; зазвичай поділяються на нюхову цибулину, яка приймає нюхові нитки, та з’єднувальну, стоншену частину – нюховий тракт. У ссавців із добре розвиненим нюхом (макросмати) нюхова частка має внутрішню порожнину, вузьким каналом вона з’єднується із бічними шлуночками. У ссавців, які постійно живуть у воді (китоподібні тощо), нюхові частки зовсім атрофовані.
У дуже багатьох ссавців кора півкуль утворює складки і звивини, розділені глибокими борознами. Ссавці малих розмірів і більш низької організації мають півкулі із гладкою поверхнею. У тварин, більших за організацією і розмірами, поверхні півкуль покрита багатьма звивинами.
Головне у філогенезі головного мозку хребетних – розвиток кінцевого мозку і особливо його плащового відділу. В амфібій з’являється закладка плаща, яка у риб тільки намічається; у рептилій уже утворюється справжня кора – архікортекс, у ссавців кінцевий мозок значно переважає решту відділів головного мозку.
Вивчення абсолютної і відносної ваги головного мозку представників різних класів хребетних показує наступне: 1) вага мозку порівняно з розмірами тіла наростає у дуже незначних розмірах. Такі велетні, як слон, мають мозок, який тільки у три – чотири рази переважає людський. 2) Тільки у небагатьох (найбільших) тварин абсолютна вага мозку більша, ніж у людини. 3) За вагою свого мозку людина значно переважає найбільш близько розміщених до неї людиноподібних мавп. Відносна вага мозку велика у приматів, а потім у тварин із незначними розмірами тіла.
Короткий нарис ембріогенезу головного мозку
Уже на ранніх стадіях розвитку мозкової трубки передній її кінець виділяється за своєю шириною – намічається зачаток головного мозку, який потім двома перехватами поділяється на три первинних мозкових пухирі: 1) передній мозок, prosencephalon; 2) середній мозок – mesencephalon; 3) ромбоподібний мозок – rhombencephalon. Дуже швидко перший і третій пухирі поділяються кожен ще на два вторинних пухирі. Перший – на кінцевий мозок, telencephalon і проміжний – diencephalon; третій поділяється на задній мозок, metencephalon і довгастий мозок, myelencephalon.
У зародків ссавців і людини зазначені відділи мозку тільки в самому початку розвитку лежать в одній площині; в подальшому у зв’язку із швидким ростом головного мозку утворюються вигини: тім’яний – у ділянці середнього мозку, мостовий – у ділянці заднього мозку, і потиличний – у ділянці довгастого мозку, на межі зі спинним мозком. Перший і третій вигини випуклістю звернені дорзально, другий – вентрально. Окремі частини головного мозку ростуть нерівномірно; цим пояснюється утворення різного роду складок, неоднаковий ріст у товщину стінок мозкових пухирів. Таким чином, головний мозок людини набуває у вищому ступені складну будову. Відповідно змінюється внутрішня порожнина головного мозку – розвиваються шлуночки (продовження центрального каналу спинного мозку).
Ромбоподібний мозок диференціюється на задній мозок, із якого виділяється перешийок ром подібного мозку, та довгастий мозок. Із перешийка, менш значного за масою відділу, розміщеного найбільше спереду, утворюються верхні ніжки мозочка і верхній мозковий парус, які безпосередньо переходять у середній мозок. В утворенні заднього і довгастого мозку головну участь беруть, як і у розвитку спинного мозку, бічні пластинки; останні, розростаючись у ділянці заднього мозку у вентральному напрямку, дають початок мосту, який спочатку являє собою вузьку поперечну смужку. Дорзально розвивається мозочок: раніше – його непарна частина – черв’як, потім – півкулі, далі з’являються борозни, які поділяють мозочок на часточки і звивини.
Стінка ромбоподібного мозку сильно потовщується – як із боків, так і на дні; тільки дах залишається у вигляді тонкої пластинки– епендими, яка у поєднанні з м’якою оболонкою головного мозку утворює судинну основу четвертого шлуночка. Остання частково атрофується, так що у подальшому четвертий шлуночок здується із субарахноїдальним простором за участю непарного середнього отвору четвертого шлуночка, apertura mediana ventriculi quarti і парного бічного, apertura lateralis ventriculi quarti. Порожнина ромбоподібного мозку перетворюється на четвертий шлуночок; спереду він сполучається із водопроводом мозку, ззаду – із центральним каналом спинного мозку і з субарахноїдальним простором.
Середній мозковий пухир розвивається найбільш просто і відстає у розвитку. Стінки його потовщуються рівномірно, порожнина перетворюється на водопровід, який з’єднує порожнину третього і четвертого шлуночків. Із дорзальної стінки походить чотиригорб’я (пластинка покрівлі), lamina tectalis; із вентральної – ніжки мозку, pedunculi cerebri.
Проміжний мозок функціонально і морфологічно пов’язаний із органом зору; на дуже ранній стадії розвитку, коли межа між кінцевим і проміжним мозком ще слабо виражена, на базальній частині бічних стінок проміжного мозку утворюються парні вирости – очні пухирі, які залишаються у зв’язку з місцем їх утворення за допомогою очних стеблин, які пізніше перетворюються у зорові нерви. Із стінок проміжного мозку найбільшої товщини досягають бічні, тут розвивають зорові горби, згір’я, які являють собою поєднання кількох сірих ядер. У зв’язку з цим порожнина проміжного мозку – третій шлуночок – перетворюється на вузьку щілину. Решта стінок залишається порівняно тонкими, у вентральній ділянці – підзгір’ї утворюється непарне випинання – лійка, infundibulum, із нижнього кінця якої походить задня (мозкова) частка гіпофіза – нейрогіпофіз. До підзгір’я відноситься також сірий горб, tuber cinereum, і сосочкові тіла – corpora mamillaria. У ділянці дорзальної стінки розвивається верхній додаток мозку – шишкоподібне тіло, corpus pineale. У більшій своїй частині дорзальна стінка залишається особливо тонкою, зберігаючи будову епендими, яка, поєднуючись із дуплікатурою м'якої оболонки, дає верхній судинний шар, tela chorioidea superior, останній бере участь в утворенні верхньої стінки третього шлуночка.
Кінцевий мозок на початку розвитку лежить спереду від решти відділів мозку і за величиною поступається їм. Але дуже швидко він у своєму рості випереджує решту мозкових пухирів і, покриваючи їх послідовно один за іншим, розповсюджується назад, утворюючи плащ мозку, pallium.
Диференціювання кінцевого мозку. Із спершу непарного утворення розвиваються два вирости – права та ліва півкулі великого мозку, тим самим порожнина кінцевого мозку, спочатку спільна, поділяється на два бічних шлуночки мозку. У міру того, як серединна щілина (майбутня поздовжня щілина мозку) між півкулями поглиблюється. Отвір, який поєднує шлуночки з порожниною другого мозкового пухиря, поділяється на два міжшлуночкових отвори, foramina interventriculariа. Ці міжшлуночкові отвори поєднують порожнину третього шлуночка із порожнинами бічних шлуночків. Стінки бічних шлуночків досягають значної товщини; порожнини шлуночків звужуються. Мозкова речовина росте не рівномірно: на поверхні півкуль утворюються численні складки – звивини мозку, які відділені одна від другої більш або менш глибокими борознами і щілинами. Більш постійні із них розвиваються раніше; однією із перших з’являється бічна борозна, sulcus lateralis; вона знаходиться на бічній поверхні півкуль і спочатку має вигляд ямки, потім її краї розростаються і прикривають дно; утворюється покришка, ямка перетворюється у щілину і комплекс звивин, які розвиваються на її дні – у так званий острівець, insula. Слідом за бічною борозною в певному порядку з’являються інші борозни. Кожна півкуля поділяється на чотири частки; відповідно до цього і порожнина бічних шлуночків поділяється на чотири частини: центральна частина і три роги шлуночка. Частки поділяються на звивини, якими покривається вся поверхня півкуль. Внаслідок збільшується і поверхня мозкової кори.
Сіра речовина (нервові клітини) кінцевого мозку розміщується не тільки у складі стінок шлуночків мозку, але і на периферії, де утворює кору півкуль, cortex cerebri. Значна маса сірої речовини розвивається на основі півкуль – ядра основи: хвостате ядро, сочевицеподібне ядро, огорожа.
Спочатку півкулі з’єднані між собою тільки у місцях їх походження із кінцевого мозку, звернені одна до іншої медіальні поверхні між собою не зв’язані. Лише значно пізніше (у зародка 7 мм довжини) виникає спільний зачаток мозкових спайок, із якого походять мозолисте тіло, склепіння і передня спайка. До похідних кінцевого мозку відносяться також нюхові частки, які у людини піддалися значній редукції.
Будова спинного мозку
Спинний мозок, medulla spinalis (грецька – mielos), являє собою у дорослої людини довгий, приблизно циліндричної форми тяж, трохи здавлений у напрямку спереду назад і вигнутий відповідно до шийного і грудного вигинів хребта. Вгорі він переходить у довгастий мозок, внизу закінчується конусом, conus medullaris.
Від зверненої вниз верхівки конусу тягнеться тонка кінцева нитка, filum terminale, яка закінчується в окісті ІІ куприкового хребця. Кінцева нитка у верхньому своєму відділі – рудимент спинного мозку.
У перші місяці внутрішньоутробного розвитку хребет і спинний мозок ростуть рівномірно; останній займає хребтовий (і крижовий) канал на всьому його протязі, а корінці усіх спинномозкових нервів, відходячи від спинного мозку під прямим кутом, направляються у відповідні міжхребцеві отвори. Із четвертого місяця розвитку плода спинний мозок починає відставати у своєму рості. Краніальний його кінець у місці переходу в довгастий мозок фіксований, тому топічні відносини спинного мозку і хребта у верхніх відділах тіла не змінюються; каудальні кінці все більше розходяться, і у новонародженого нижній кінець спинного мозку знаходиться на рівні ІІІ поперекового хребця, у дорослого – на рівні І поперекового хребця. Внаслідок цього, корінці нижніх поперекових і всіх крижових нервів набувають косий (а найбільш каудально розміщені – майже вниз) напряму і, перед тим, як досягти відповідних міжхребцевих отворів, проходять всередині хребтового (і крижового) каналу на більшому або меншому протязі. Комплекс корінців (передніх і задніх) чотирьох нижніх поперекових, п’яти крижових і одного куприкового нервів має назву кінського хвоста, cauda equina; до його складу також входить і кінцева нитка. Верхня межа спинного мозку відповідає рівню виходу корінців І спинномозкового нерва – співпадає із верхнім краєм першого шийного хребця, нижні у дорослого чоловіка опускається до І поперекового, у жінки – заходить у ділянку ІІ поперекового хребця. Довжина його в середньому дорівнює 45 см, вага – 30 г. Товщина спинного мозку по всій довжині неоднакова, є два потовщення: шийне, intumescentia cervicalis, тягнеться від ІІ шийного до ІІ грудного хребця, краще всього виражене на рівні V – VI шийних хребців; поперекове, intumescentia lumbalis, починається на рівні Х грудного хребця, найбільшу товщину має на ХІІ грудному хребці, потім переходить у мозковий конус.
Потовщення розміщені там, де від спинного мозку відходять найтовстіші нарви (до кінцівок); утворення потовщень пов’язане зі збільшенням кількості нервових клітин і волокон у відповідних відділах спинного мозку.
Спинний мозок складається із центрально розміщеної сірої речовини, substantia grisea, і білої речовини, substantia alba, яка її оточує. Він на всю довжину поділений на дві симетричні половини – праву та ліву за допомогою двох борозен, які проходять у серединній площині; передня, fissura mediana anterior і задня, fissura mediana posterior. До останньої примикає гліальна перегородка, яка простягається у серединній площині на всю товщину заднього відділу білої речовини до задньої сірої спайки.
Окрім вище зазначених борозен, є ще дві бічні (парні): бічна передня та задня, права та ліва борозни, sulcus lateralis anterior et posterior dexter et sinister. Ці борозни, особливо задня бічна, виражені слабко, але ними користуються для визначення розподілу білої речовини спинного мозку на канатики – три з кожного боку: передній канатик, funiculis anterior, задній канатик, funiculis posterior, бічний канатик, funiculis lateralis.
Вздовж передньо-бічних борозенок спинного мозку виходять із двох боків передні (вентральні) корінці, вздовж задньо-бічних виходять задні (дорзальні) корінці спинномозкових нервів. Передній корінець, radix anterior, складається із рухових (еферентних) нервових волокон; задній корінець, radix posterior, складається із чутливих (аферентних) провідників. Поблизу спинного мозку (більшою частиною у міжхребцевих отворах), з обох боків передні та задні корінці з’єднуються і утворюють стовбур спинномозкового нерва, truncus nervi spinalis. Він має змішаний характер (руховий і чутливий). Стовбури виходять із спинномозкового каналу через міжхребцеві отвори. Кожен задній корінець у місці свого з’єднання із переднім має потовщення – спинномозковий вузол, ganglion spinale. Вузли містять несправжньооднополюсні нейрони. Їх єдиний відросток Т–подібно ділиться на два більш тонких; один із них іде на периферію, де закінчується певним нервовим закінченням; другий направляється до центру (до спинного мозку). Сукупність центральних відростків утворює задній корінець. Усього спинномозкових нервів 31 пара. Ділянка сірої і білої речовини, яка відповідає кожній парі спинномозкових нервів, називається спинномозковим сегментом. Таким чином, спинний мозок складається із 31 сегмента: 8 шийних, 12 грудних, 5 поперекових, 5 крижових та 1 куприкового.
Сіра речовина, substantia grisea, присутня на всьому протязі спинного мозку. Вона знаходиться всередині спинного мозку і оточена з усіх боків білою речовиною, substantia alba. Сіра речовина утворює дві вертикальні колони, розміщені у кожній половинці спинного мозку. Всередині його закладений центральний канал, canalis centralis, який містить спинномозкову рідину. Центральний канал є залишком порожнини первинної нервової трубки, тому вверху він сполучається із четвертим шлуночком головного мозку, а у ділянці мозкового конусу закінчується розширенням – кінцевим шлуночком, ventriculus terminalis. Сіра речовина, яка оточує центральний канал, має назву проміжної, substantia intermedia centralis. У кожній колоні сірої речовини є два стовпи: передній, columna anterior та задній, columna posterior.
На поперечних зрізах спинного мозку ці стовпи мають вигляд рогів: переднього, розширеного, cornu anterius, та заднього, загостреного, cornu posterius. Тому загальний вигляд сірої речовини на фоні білої нагадує літеру “Н”.
Нервова система являє собою складний комплекс багатьох нейронів, з’єднаних між собою відростками, по яких передається збудження. Шлях, по якому проходить збудження і відповідь на нього, називається рефлекторною дугою.
Рефлекторна дуга
У простому вигляді рефлекторна дуга складається тільки із трьох нейронів – чутливого, вставного та рухового. Прикладом може служити сухожилковий колінний рефлекс. При подразненні (наприклад, постукуванні молоточком) пропріоцепторів, розміщених у товщі зв’язки надколінка (яка є сухожилком чотириголового м’яза стегна
Дата добавления: 2014-12-03; просмотров: 510; Мы поможем в написании вашей работы!; Нарушение авторских прав |