КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Функциональная структура КИС
Закон силы раздражения. Качественные и количественные изменения процессов, протекающих в организме, соответственно отображают качественные и количественные особенности действующих на него раздражителей и способа их действия на организм, т. е. раздражения.
Наименьшая сила раздражителя, которая вызывает минимальное возбуждение, называется порогом раздражения. Так как порог раздражения характеризует возбудимость, то он является вместе с тем и порогом возбудимости. Чем больше возбудимость, тем все более уменьшается порог раздражения, и, наоборот, чем меньше возбудимость, тем больше сила раздражения, которая вызывает наименьшее возбуждение. Порог возбудимости определяется на нервно-мышечном препарате по силе постоянного электрического тока, необходимого для получения едва заметного сокращения мышцы.
Чем больше сила раздражения, тем до определенного предела больше возбуждение и, следовательно, ответная реакция возбуждаемой ткани.
Сила раздражения меньше пороговой называется подпороговой, а больше пороговой — надпороговой. Наименьшая сила раздражения, вызывающая наибольшую ответную реакцию ткани, называется максимальной. Разные нарастающие величины силы раздражения, расположенные между пороговой и максимальной, называются субмаксимальными, а больше максимальной — сверх- максимальными.
Порог возбудимости зависит от свойств возбудимой ткани, от ее физиологического состояния в момент нанесения раздражения, от способа и продолжительности раздражения и от крутизны нарастания силы раздражения.
Закон градиента раздражения (аккомодация). В 1848 г. Дюбуа- Реймон обнаружил, что если через нерв или любую другую ткань проходит постоянный электрическии ток пороговой силы и сила этого тока на протяжении значительного отрезка времени не меняется, то такой ток при своем прохождении не вызывает возбуждения ткани. Возбуждение возникает только в том случае, есди сила электрического раздражителя быстро нарастает или убывает. При очень медленном нарастании силы тока раздражения нет. Закон Дюбуа-Реймона относится не только к действию электрического тока, но и к действию любого другого раздражителя. Это — закон градиента. Градиентом раздражения обозначается быстрота возрастания силы раздражения. Чем больше ее увеличение в каждую последующую единицу времени, тем до определенного предела больше реакция живой ткани на это раздражение. Быстрота нарастания возбуждения зависит от градиента раздражения. Возбуждение возрастает тем медленнее, чем меньше градиент раздражения. : -
 Порог возбудимости значительно повышается при медленном нарастании раздражения. Можно предполагать, что живая ткань противодействует внешнему раздражению. Например, если быстро ударить по нерву, очень быстро его охладить или нагреть при силе раздражения выше пороговой, то возникает возбуждение. Если же.медленно надавливать на нерв, медленно его охлаждать или нагревать, то возбуждение не вызывается. Синусоидальный переменный электрический ток низкой частоты не вызывает возбуждение, так как скорость его изменения слишком мала. Следовательно, при медленном нарастании раздражения возникает приспособление, адаптация раздражаемой ткани к раздражителю (И. С. Беритов, Д. С. Воронцов). Это приспособление называется аккомодацией.
Порог возбудимости значительно повышается при медленном нарастании раздражения. Можно предполагать, что живая ткань противодействует внешнему раздражению. Например, если быстро ударить по нерву, очень быстро его охладить или нагреть при силе раздражения выше пороговой, то возникает возбуждение. Если же.медленно надавливать на нерв, медленно его охлаждать или нагревать, то возбуждение не вызывается. Синусоидальный переменный электрический ток низкой частоты не вызывает возбуждение, так как скорость его изменения слишком мала. Следовательно, при медленном нарастании раздражения возникает приспособление, адаптация раздражаемой ткани к раздражителю (И. С. Беритов, Д. С. Воронцов). Это приспособление называется аккомодацией.
Чем быстрее нарастает сила раздражения, тем до определенного предела сильнее возбуждение, и наоборот. Показатель скорости аккомодации — наименьшая крутизна нарастания силы раздражения, при которой оно еще вызывает возбуждение. Это пороговый градиент аккомодации.
У двигательных нервов аккомодация значительно больше, чем у чувствительных. Самая малая аккомодация у тканей, обладающих автоматизмом (сердечной мышцы, гладкой мускулатуры пищеварительного канала и других органов).
Закон гиперболы. Для получения возбуждения необходимо некоторое минимальное время раздражения постоянным электрическим током. Существует определенная зависимость между силой раздражающего постоянного электрического тока и временем раздражения, необходимым для возникновения возбуждения, или латентным периодом. Эта зависимость выражается кривой силы — времени, имеющей вид равносторонней гиперболы (Гоор- вег, 1892, Вейс, 1901) (рис. 95).
Закон гиперболы: каждому минимальному промежутку времени раздражения соответствует минимальная сила постоянного тока, при которой получается возбуждение, и наоборот. В современной физиологии существуют электронные приборы, позволяющие раздражать ткань в течение тысячных и меньше долей секунды или в микроинтервалах времени (0,001 с сокращенно обозначается буквой СУ — сигма).
Чем сильнее ток, тем меньше продолжительность его действия, необходимая для получения возбуждения, и наоборот.
Полярный закон Пфлюгера. Пфлюгер (1859) установил, что при раздражении постоянным электрическим током возбуждение возникает в момент его замыкания или при возрастании его силы в области приложения к раздражаемой ткани отрицательного полюса — катода, откуда оно распространяется вдоль по нерву или мышце. В момент размыкания тока или при его ослаблении возбуждение возникает в области приложения положительного полюса — анода. При одной и той же силе тока возбуждение больше при замыкании в области катода, чем при размыкании в области анода. При раздражении нервно-мышечного препарата постоянным электрическим током получаются различные результаты в зависимости от его силы и направления. Различают восходящее направление тока, при котором ближе к мышце расположен анод, и нисходящее — если ближе к мышце расположен катод. Явления электротона и периэлектротона. При замыкании и прохождении постоянного тока через нерв или мышну изменяются физиологические и физико-химические свойства на полюсах.
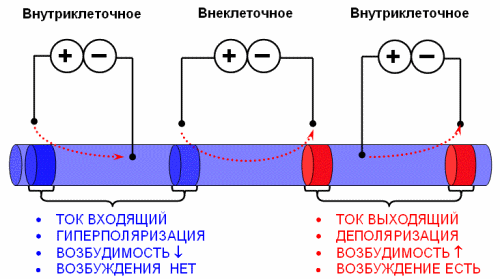 При прохождении постоянного тока в области приложения катода возбудимость временно повышается, а в области приложения анода возбудимость временно понижается (рис. 96). Даже слабые и кратковременные токи вслед за повышением, возбудимости вызывают понижение возбудимости в области действия катода. Особенно отчетливо выступает это последующее понижение возбудимости в этой области при действии относительно сильных и продолжительных токов — катодическая депрессия (Б. Ф. Вериг о, ! 1888). Катодическая депрессия может прекратить проведение нервных импульсов (Д.. С. Воронцов, 1937). Она исчезает через
При прохождении постоянного тока в области приложения катода возбудимость временно повышается, а в области приложения анода возбудимость временно понижается (рис. 96). Даже слабые и кратковременные токи вслед за повышением, возбудимости вызывают понижение возбудимости в области действия катода. Особенно отчетливо выступает это последующее понижение возбудимости в этой области при действии относительно сильных и продолжительных токов — катодическая депрессия (Б. Ф. Вериг о, ! 1888). Катодическая депрессия может прекратить проведение нервных импульсов (Д.. С. Воронцов, 1937). Она исчезает через
— 8 мс после выключения постоянного тока.
В области действия катода при замыкании увеличивается скорость проведения возбуждения, а в области действия анода она уменьшается. В области действия катода уменьшается высота волны возбуждения и увеличивается ее продолжительность, а в области действия анода, наоборот, высота увеличивается, а продолжительность ее уменьшается. Продолжительность полной не- |Возбудимости в области действия катода увеличивается, а анода — уменьшается. Поэтому лабильность в области действия катода уменьшается, а в области действия анода увеличивается.
Эти изменения физиологических свойств нерва в области действия катода обозначаются как катэлектротон, а в области действия анода — как анэлектротон. Изменения физиологических свойств нерва происходят не только в месте приложения полюсов постоянного тока, но и на некотором расстоянии от них. На расстоянии около 2 см за пределами катода возбудимость нерва понижается, а за пределами анода она повышается. Этот факт открыл Н. Я. Перна (1914) и обозначил его как периэлектротон.
Следовательно, в периферических нервах не только распространяются волны возбуждения, но при появлении очага возбуждения на некотором расстоянии от него возникают и устанавливаются вдоль всего нерва участки повышенной и пониженной возбудимости в виде стационарной волны. Таким образом, в периферических нервах имеется двойная нервная сигнализация: импульсная и тоническая. Некоторые авторы отрицают существование периэлектротона (Д. С. Воронцов, 1961). «
В пунктах приложения полюсов постоянного тока увеличивается количество ацетилхолина в области действия катода и уменьшается в области действия анода, относительно увеличивается содержание ионов калия в области действия катода и ионов кальция в области действия анода, увеличивается проницаемость белковых мембран в области действия катода и уменьшается их проницаемость в области действия анода.
 Изменения возбудимости нервов при действии постоянного тока наблюдаются и у человека. Электрод с малой поверхностью-,или дифферентный, прикладывают к раздражаемой области нерва, а электрод с большой поверхностью, или индифферентный, накладывают на отдаленную часть тела. При таком униполярном способе раздражения действие тока проявляется только вблизи дифферентного электрода. В зависимости от силы тока получают различные результаты.
Изменения возбудимости нервов при действии постоянного тока наблюдаются и у человека. Электрод с малой поверхностью-,или дифферентный, прикладывают к раздражаемой области нерва, а электрод с большой поверхностью, или индифферентный, накладывают на отдаленную часть тела. При таком униполярном способе раздражения действие тока проявляется только вблизи дифферентного электрода. В зависимости от силы тока получают различные результаты.
При слабой силе постоянного тока раздражение в области анода является подпороговым. Поэтому независимо 'от направления тока сокращение получается только в области катода, так как возбуждение на этом полюсе больше, чем на аноде. При средней силе тока раздражение в области анода достигает порога. Поэтому независимо от направления тока сокращения получаются и в области катода и в области анода.
При сильном восходящем токе возбуждение возникает в области катода при замыкании, но оно не может достигнуть мышцы, так как по пути возникает анэлектротон (резкое понижение возбудимости и проводимости), поэтому сокращение получается только при размыкании. При сильном нисходящем токе замыкание вызывает сокращение мышцы, а при размыкании сокращения нет. Это отсутствие сокращения зависит от того, что в момент размыкания в области катода возбудимость и проводимость резко понижаются и возбуждение, возникающее на аноде, не проводится к мышце (табл. 9).
Возбудимость и ее динамика. Мера возбудимости живой клетки определяется по двум показателям: 1) по наименьшей пороговой силе (интенсивности) раздражения, вызывающего возбуждение, которая называется порогом возбудимости (см. с. 344), и 2) по наименьшему времени действия раздражителя определенной силы (интенсивности).
Возбудимость каждой живой ткани изменяется в зависимости от условий и от ее физиологического состояния: например, при постепенном охлаждении, при смещении реакции крови в сторону кислотности она снижается, а при постепенном повышении температуры до 40°С и смещении реакции крови в сторону щелочности повышается.
У животных с постоянной температурой тела исходный уровень возбудимости, характеризующий данную живую ткань, наблюдается при отсутствии утомления, при нормальной температуре тела и нормальной реакции крови.
Лабильность. Н. Е. Введенский развил представление о лабильности, или функциональной подвижности ткани (1892). Он определял физиологическую лабильность как скорость, с которой данная живая ткань успевает закончить полный период отдельного возбуждения во времени.
А. А. Ухтомский считал, что мера лабильности — это наибольшее «число отдельных законченных периодов возбуждения, которое субстрат может вместить в единицу времени».
Физиологическая лабильность — основное свойство живой ткани, определяющее ее функциональное состояние. Она характеризует изменения физиологического состояния живой ткани не при одиночной волне возбуждения, а при взаимодействии целого ряда волн возбуждения, возникающих в определенном ритме, — ансамбля возбуждений. От лабильности зависит, будет ли живая ткань отвечать волной возбуждения на каждый импульс ритмического раздражения или она будет трансформировать частый ритм раздражения в более редкий, или такое трансформирование пре-вратится в торможение, а торможение снова превратится в возбуждение.
Чем больше возрастает частота импульсов раздражения, тем чаще становится ритм волн возбуждения. Максимальный ритм раздражения вызывает максимальный ритм возбуждения, который отличается большой неустойчивостью. Электрофизиологиче- скими исследованиями установлено, что каждая живая ткань способна воспроизводить синхронно, т. е. соответственно ритму раздражения без трансформации торможения или утомления, характерный для нее оптимальный ритм раздражения.
Максимальный ритм синхронизированного ответа на раздражение для одиночных двигательных нервных волокон лягушки около 300 в 1 с, оптимальный — 75 (реже 50) — 150 в 1 с, для мышечных волокон максимальный — 150 (реже 200) в 1 с, оптимальный — 20 — 50 в 1 с.
Максимальный ритм проведения импульсов в двигательных нервах теплокровных больше 1000 в 1 с, а в нервных центрах—200—400 в 1 с. Н. Е. Введенский установил, что сами импульсы возбуждения способны изменять лабильность раздражаемой ткани, повышать ее и понижать.
Физиологическая лабильность данной ткани зависит от силы и частоты импульсов возбуждения, поступающих к ней из центральной нервной системы, и от нервно-гуморальных влияний. Имеется зависимость между физиологической лабильностью и возбудимостью. Возбудимость ткани наивысшая при среднем, относительно невысоком уровне физиологической лабильности. Лабильность ткани тем больше, чем меньше времени необходимо для возникновения возбуждения при раздражении. Лабильность тем меньще, чем медленнее реагируют ткани на раздражение. Лабильность определяет не только минимальное время, необходимое для возникновения возбуждения, но и все время, необходимое для протекания возбуждения и для восстановления способности ткани давать новые, последующие импульсы возбуждения. Условия, понижающие жизнеспособность ткани (холод, нагревание, сильный электрический ток, механическое давление, наркотики, солевые растворы и т. п.), уменьшают лабильность измененного (альтерированного) этими воздействиями участка нерва. Это уменьшение лабильности обусловлено тем, что под влиянием указанных воздействий замедляются восстановительные процессы.
Различные i рупиы нервных волокон обладают разной лабильностью. Лабильность одних и тех же нервных волокон колеблется в зависимости от их физиологического состояния.
Усвоение ритма. Самый частый ритм пороговых и надпорого- вых раздражений, на который данная возбудимая ткань отвечаем таким же частым ритмом волн возбуждения, отражает ее функционально^ состояние или ее лабильность во время деятельности.
А. А. Ухтомский создал представление об усвоении ритма (1928), согласно которому лабильность меняется все время в связи с деятельностью. Лабильность во время раздражения может повышаться или понижаться, что выражается в увеличении или уменьшении предельного ритма возбуждения. Это изменение лабильности вызывается тем, что сами импульсы, возбуждения способны изменить функциональное состояние возбуждаемой ткани. После действия каждого раздражающего импульса лабильность изменяется двухфазно: вначале она повышается, а затем падает. Лабильность зависит от силы и частоты падающих на ткань импульсов и от обмена веществ в ткани.
Под влиянием работы лабильность повышается, что приводит к усвоению более высокого ритма, чем в начале работы. Усвоение ритма особенно отчетливо выступает на фоне повышенной возбудимости. Оно продолжается некоторое время после прекращения работы.
Повышение физиологической лабильности в связи с деятельностью, которое проявляется в том, что возбудимая ткань отвечает более высоким ритмом возбуждения по сравнению с исходным ритмом, называется усвоением, ритма. Усвоение ритма зависит от текущих изменений обмена веществ в ткани во время се деятельности. После короткого раздражения мышцы ее лабильность повышается в течение нескольких минут, что можно объяснить действием продуктов обмена веществ.
Реобаза и хронаксия. Зависимость латентного периода возникновения возбуждения от силы раздражающего электрического тока представлена кривой сила — время (рис. 97).
При раздражении поверхностной мембраны клетки через нее проходит минимальное количество электричества, измеряемое произведением силы тока на время его действия. Каждой продол жительности времени раздражения соответствует минимальная сила тока,, впервые вызывающая воз-буждение в раздражаемой ткани, Следовательно, возникновение возбуждения зависит не толь-, ко от силы раздражения, но t-и от продолжительности его действия. Эта закономерность сначала была открыта на медленно реагирующей мышце пресноводного моллюска анодонты, д затем на гладкой мышце кролика. С изобретением приборов, позволяющих дозировать раздражение “"в течение долей миллисекунды, она была обнаружена и на нервах и мышцах человека и возбудимых тканях позвоночных животных.
Минимальное время действия на поверхностную мембрану клетки прямоугольного толчка постоянного электрического тока, вызывающего изменение разности потенциалов при возникнове-нии возбуждения, определяется ее сопротивлением и емкостью. Минимальная, или пороговая, сила тока, вызывающая изменение разности потенциалов мембраны, называется реобазой, а минимальное время действия реобазы — полезным временем. Определить полезное время (точку 1) трудно, так как соседние точки мало от нее отличаются, вправо и влево „от точки 1 кривая изменяется незначительно. Поэтому французский физиолог JI. Лапик (1909) предложил вместо полезного времени определять минимальное время действия тока в две реобазы, которое обозначается как хронаксия. Точка 2 (хронаксия) легко и точно определяется, так как изменения кривой вправо и влево от нее значительны. Время раздражения вправо от точки 1 не имеет значения. Следовательно, когда кривая идет параллельно оси абсцисс, действует закон Дюбуа-Реймона. Когда продолжительность раздра- жен'ия становится меньше полезного времени, сила тока, вызывающая возбуждение, круто возрастает влево от точки 1. На кривой видно, что чем короче время раздражения, тем больше должна быть сила раздражающего тока (точки 2 и 3). Затем наступает такой момент, когда кривая идет параллельно оси ординат (точка 4). Теперь уже сила тока при небольшом сдвиге влево не имеет значения, так как скорость ее изменения слишком мала. Поэтому переменный ток очень высокой частоты не вызывает возбуждения, вследствие того что его полупериод очень короток и недостаточен для изменения потенциала мембраны.
Влево от этой точки дальнейшее уменьшение времени приводит к отсутствию возбуждения даже при очень больших напряжениях тока. При достаточно больших промежутках времени действия постоянного электрического тока пороговая сила раздражения зависит не только от продолжительности его действия, но и от аккомодации.
Кривая сила — время представляет собой равностороннюю гиперболу, которая имеет формулу
= ~+b, t
где г — пороговая сила тока; t — пороговая длительность раздражения; а — константа, характеризующая постоянное время раздражения с момента, когда кривая переходит в прямую линию, идущую параллельно оси ординат; b — константа, соответствующая силе раздражения при постоянной его длительности, когда кривая переходит в линию, идущую параллельно оси абсцисс.
Если пороговая сила тока равна 2Ь, т. е. двум реобазам, то формула превращается в Ь — следовательно,'
При Ь условно принятым за единицу t = а.
Следовательно, t — наименьшее время раздражения при силе тока, равной двум реобазам, или хронаксия.
Для любой возбудимой ткани кривая, выражающая соотношение между силой постоянного тока и временем его действия, имеет одинаковую форму равносторонней гиперболы. Но минимальное время, в течение которого вызывается возбуждение при действии тока в две реобазы, для каждой ткани различно и определяется ее функциональной подвижностью.
Понятие лабильности включает хронаксию. Хронаксия — это только показатель минимального времени, в течение которого должен действовать раздражитель определенной силы, для того чтобы возникло возбуждение, а лабильность характеризует все время, необходимое для развития и прекращения возбуждения. Хронаксия тем больше, чем медленнее реагируют живые ткани па раздражение. Хронаксия скелетных мышц человека колеблется от 0,1 до 0,7 мс. Хронаксия сгибателей у человека в 1,5—2 раза меньше хронаксии разгибателей. После перерезки и перерождения двигательного нерва хронаксия мышцы удлиняется приблизительно в 10 раз. Хронаксия тем меньше, чем толще мышечные волокна. При ухудшении функционального состояния хронаксия сначала уменьшается, а затем возрастает. Для того чтобы возбуждение могло перейти с одной живой ткани на другую, например с нерва на мышцу, необходимо, чтобы у обеих тканей была приблизительно одинаковая хронаксия (изохронизм). Однако в некоторых случаях возбуждение передается с нерва на мышцу и при отсутствии изохронизма, когда имеется значительная разница в хронаксии. Это расхождение в хронаксии обозначается как гете- рохронизм.
Раздражения разных внутренних органов вызывают закономерные изменения хронаксии скелетных мышц (С. И. Гальперин, М. Р. Могендович, 1941; 1942).
Хронаксию определяют не только в экспериментальных физиологических исследованиях, но также и в клинике, так как она один из показателей функциональной подвижности ткани или органа. Хронаксия разных групп нервных волокон неодинакова. У животных с постоянной температурой тела хронаксия безмякотных волокон равна десяткам миллисекунд. Хронаксия нервов изменяется в разных участках.
Сравнительно-физиологические и возрастные особенности возбудимости нервов и мышц. Н. Е. Введенский (1886) предполагал, что чем выше уровень развития живой ткани в филогенезе, тем быстрее возникает в ней возбуждение при пороговом раздражении. Это подтвердилось в опытах с определением продолжительности полезного времени.
В процессе эволюции ткани становятся все более быстрореаги- рующими. Чем выше развитие ткани, тем короче ее полезное время. Например, полезное время гладкой мышцы желудка лягушки равно нескольким секундам, мышцы сердца моллюска — десятым долям секунды, мышцы ноги моллюска — сотым долям секунды, двигательного нерва лягушки — тысячным долям секунды, двигательного нерва животных с постоянной температурой тела — десятитысячным долям секунды.
Возбудимость скелетной мышцы значительно ниже в тех ее участках, где нет мионевральных аппаратов.
Возбудимость нервных волокон зависит от их толщины: чем они толще, тем выше возбудимость. Наибольшая возбудимость у волокон группы А, наименьшая — у волокон группы С, средняя — у волокон группы .б.Если принять пороговую величину одиночного удара индукционного тока для волокон группы А за единицу, то для волокон группы В она равна 2,4, а для волокон группы С — 15,7.
Чем выше организация животного, тем больше у него период физиологического покоя, так как у него больше лабильность и, следовательно, быстрее происходит возврат в состояние покоя (А. А. Ухтомский, 1937).
У беспозвоночных хронаксия значительно больше, чем у позвоночных. В двигательных нервах, иннервирующих гладкие мышцы, и Bi гладких мышцах позвоночных она в десятки раз больше, чем в двигательных нервах скелетных мышц и в скелетных мышцах.
В онтогенезе хронаксия резко уменьшается. Например, у пятидневного цыпленка хронаксия одной из мышц в 60 раз больше, чем той же мышцы взрослой курицы. У ребенка хронаксия скелет ных мышц к 1 году уменьшается в 3 — 4 раза. Хронаксия двуглавой мышцы плеча у ребенка 4 — 6 лет равна 0,17 мс, 6 — 8 лет — 0,11 мс,
—10 лет — 0,10 мс. В течение нескольких часов после рождения она в 6 раз больше, чем у взрослых.
С возрастом хронаксия становится более постоянной.
На ранних стадиях онтогенеза отсутствуют резкие различия лабильности антагонистических мышц, характерные для взрослых организмов.
Уменьшение хронаксии в фило- и онтогенезе обусловлено ускорением обмена веществ в нервах и мышцах и укорочением времени протекания возбуждения. Чем меньше хронаксия нерва или мышцы, тем больше скорость распространения в них возбуждения.
Функциональная структура КИС
Дата добавления: 2015-01-05; просмотров: 638; Мы поможем в написании вашей работы!; Нарушение авторских прав |