КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
КЛІТИННА ОБОЛОНКА. ЦИТОПЛАЗМА
Клітинна оболонка (plasmolemma).Воснові будови плазмолеми лежить елементарна біологічна мембрана. Структуру останньої описує рідинно-мозаїчна модель Сінгер-Нікольсона. Суть її така: молекули фосфоліпідів, контактуючи своїми гідрофобними кінцями і відштовхуючись гідрофільними, утворюють суцільний подвійний ліпідний шар, у який частково або повністю втоплені молекули білків (переважно глікопротеїнів). Молекули білків, які пронизують усю товщу біліпідного шару або у значній мірі втоплені в нього,— це так звані інтегральні білки; ті ж білки, які розміщені лише на поверхні ліпідів, називаються периферійними, або адсорбованими (рис. 1.2). Положення білкових молекул не є жорстко лімітованим — залежно від функціонального стану клітини може відбуватися їхнє взаємне переміщення у площині біліпідного шару. Ця мінливість і подібна до мозаїки топографія макромолекулярних комплексів поверхні клітини дала назву рідинно-мозаїчній моделі біологічної мембрани. Лабільність (плинність) структур плазмолеми залежить від вмісту у її складі молекул холестерину: чим вищий вміст холестерину, тим легше переміщаються макромолекулярні білкові комплекси у біліпідному шарі. Серед білків плазмолеми існує певна спеціалізація: є структурні, ферментні, транспортні, рецепторні молекули. Однією з важливих умов нормального функціонування біологічної мембрани є збереження принципу замкнутості (відсутності розривів) біліпідного шару.
Вуглеводні компоненти молекул глікопротеїнів та гліколіпідів плазмолеми, виступаючи над зовнішньою поверхнею клітинної мембрани, формують так звану надмембранну зону, або глікокалікс. Олігосахаридні ланцюги глікокаліксу служать своєрідною «візитною карткою» клітини. За їхньою участю здійснюється взаєморозпізнавання клітин та взаємодія з мікрооточенням. Кожному різновиду клітин притаманна особлива послідовність моносахаридних залишків у складі поверхневих олігосахаридних ланцюгів глікополімерів, свій унікальний набір і цитотопографія вуглеводних детермінант.
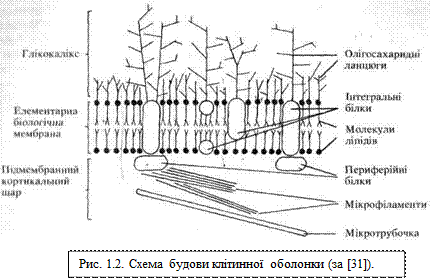 З боку внутрішнього вмісту клітини з мембраною контактує так звана внутрішня (підмембранна) пластинка, або кортикальний шар плазмолеми. Це найбільш в'язка частина цитоплазми, багата мікрофіламентами і мікротрубочками, які утворюють високоорганізовану сітку і за участю яких здійснюються, зокрема, переміщення інтегральних білків плазмолеми, забезпечуються цитоскелетні та локомоторні функції клітини, реалізуються процеси екзоцитозу. Товщина плазмолеми становить близько 10 нм (1 нм= 10-9 м).
З боку внутрішнього вмісту клітини з мембраною контактує так звана внутрішня (підмембранна) пластинка, або кортикальний шар плазмолеми. Це найбільш в'язка частина цитоплазми, багата мікрофіламентами і мікротрубочками, які утворюють високоорганізовану сітку і за участю яких здійснюються, зокрема, переміщення інтегральних білків плазмолеми, забезпечуються цитоскелетні та локомоторні функції клітини, реалізуються процеси екзоцитозу. Товщина плазмолеми становить близько 10 нм (1 нм= 10-9 м).
Отже, до складу клітинної оболонки входять зовнішня пластинка — глікокалікс, внутрішня пластинка — підмембранна, або кортикальна зона і проміжна пластинка — власно біологічна мембрана, для описання властивостей якої служить рідинно-мозаїчна модель.
Функції клітинної оболонки.До основних функцій плазмолеми слід віднести розмежовування внутрішнього вмісту клітини від її мікрооточення, транспорт метаболітів, у тому числі забезпечення асиметрії концентрації іонів натрію і калію в клітині і за її межами, рецепцію сигналів з боку зовнішнього середовища, забезпечення взаєморозпізнавання і взаємодії клітин з утворенням міжклітинних контактів різного ступеня складності, а також формування характерної структури клітинної поверхні.
Розмежування і транспорт— дві взаємопротилежні і взаємодоповнюючі одна одну функції плазмолеми. Завдяки розмежуванню з зовнішнім середовищем клітина зберігає свою індивідуальність, завдяки транспорту речовин може жити і функціонувати. Обидва ці процеси спрямовані на підтримання постійних характеристик внутрішнього середовища (гомеостазу) клітини. Транспорт із зовнішнього середовища всередину клітини (поглинання речовин) називається ендоцитозом, транспорт у протилежному напрямі (виведення речовин) — екзоцитозом. Невеликі молекули можуть потрапляти з зовнішнього середовища всередину клітини або шляхом дифузії (пасивний транспорт), або за участю особливих ферментів — пермеаз плазмолеми (активний транспорт).
Великі молекули та скупчення молекул поглинаються клітиною шляхом обволікання їх певною ділянкою плазмо леми з наступним втягуванням (інтернілізацією) утвореного мішечка всередину цитоплазми. Процес поглинання таким способом твердих частинок носить назву фагоцитозу, поглинання частинок рідини — п і н о ц и т о з у. Поглинуті частинки, як правило, розщеплюються і їхні хімічні складники засвоюються клітиною. Однак можливий і такий варіант, коли частинка поглинається однією поверхнею клітини, в оточенні біомембрани проходить через цитоплазму і виводиться без змін на протилежній поверхні клітини. Таке явище носить назву цитопемпсису.
Виведення клітиною продуктів життєдіяльності за межі цитоплазми (екзоцитоз) поділяється на ряд різновидів: секрецію, екскрецію, рекрецію, клазматоз. Секреція — це виділення клітиною продуктів її синтетичної діяльності, які необхідні для нормального функціонування органів та систем організму. Екскреція — виділення токсичних або шкідливих продуктів метаболізму, які підлягають виведенню за межі організму. Р е к р е ц і я — це видалення з клітин речовин, які не змінюють своєї хімічної структури у процесі внутрішньоклітинного метаболізму (вода, мінеральні солі). К л а з м а т о з — видалення за межі клітини окремих її структурних компонентів.
Механізми екзоцитозу переважно носять характер прямо протилежний до фаго- і піноцитозу. Наприклад, продукти синтетичної діяльності клітини нагромаджуються у вигляді оточених біомембраною скупчень (у складі мішечків і пухирців комплексу Гольджі), поступово зміщуються від центральних ділянок цитоплазми до периферії, після чого біомембрана мішечка включається у плазмолему, а вміст мішечка переходить у міжклітинний простір або зовнішнє середовище. Можливий також шлях виведення речовин у складі пухирців (без порушення цілості їх біомембрани), що носить назву секреції за мерокриновим типом. Коли ж багато секреторних пухирців нагромаджується у ділянці апікального полюсу клітини і вони виділяються з відривом апікального полюсу, то такий спосіб секреторної діяльності носить назву апокринового. При голокриновому способі секреції цитоплазма поступово заповнюється продуктами синтетичної діяльності й клітина перероджується в оточену плазмолемою крапельку секрету.
Примембранний метаболізм пов'язаний з присутністю на поверхні плазмолеми деяких видів клітин особливих ферментних систем, здатних розщеплювати контактуючі з ними біополімери. Такі процеси характерні, зокрема, для клітин внутрішнього вистелення тонкої кишки, на поверхні яких здійснюється так зване мембранне травлення.
Роль плазмолеми в механізмах клітинної рецепції. Сприйняття клітиною хімічних сигналів від її мікрооточення здійснюється переважно за участю спеціальних, так званих рецепторних білків плазмолеми. Для реалізації своєї дії на клітину біологічно активна речовина (наприклад, гормон), повинна зв'язатися з відповідним білком плазмолеми, причому визначальна роль у забезпеченні вибірковості (специфічності) цієї взаємодії належить структурі вуглеводного компонента (олігосахаридних ланцюгів) рецепторного білка. Один з можливих шляхів подальшої передачі інформації всередину клітини лежить через аденілатциклазну систему. Наприклад, зв'язування рецептора з відповідною хімічною речовиною зумовлює активацію розміщеного на внутрішній поверхні плазмолеми ферменту аденілатциклази, який у свою чергу стимулює утворення циклічного аденозинмонофосфату (цАМФ). Останній є універсальним активатором ряду цитоплазматичних ферментних систем, за участю яких клітина реагує на подразнення. Слід зазначити, що складні процеси рецепції — основа взаєморозпізнавання клітин, і тому є кардинально необхідною умовою існування багатоклітинних організмів, оскільки служать командою для гальмування розмноження і руху клітин. Зауважимо, що саме втрата самоконтролю клітин за ходом росту і розмноження (відсутність контактного гальмування розмноження) — один з основних проявів злоякісного переродження. Міжклітинні контакти. Клітинній оболонці, зокрема вуглеводним детермінантам її глікокаліксу, належить визначальна роль і в утворенні стійких контактів між клітинами. Найпростіша форма міжклітинного зв'язку носить назву адгезії (злипання). Останнім часом показана вирішальна роль специфічних білкових молекул — лектинів, кадгеринів і молекул клітинної адгезії, так званих САМ (від англ. Cell Adhesia Molecules) в утворенні багатоклітинних конгломератів. Молекули лектинів, зокрема, здатні вибірково «впізнавати» вуглеводні детермінанти на поверхні сусідніх клітин і забезпечувати утворення стійких міжклітинних містків. Відстань між плазмолемами суміжних клітин у зоні простого контакту становить близько 10...20 нм.
 У процесі еволюційного розвитку багатоклітинних систем значно ускладнились форми і види міжклітинних контактів, що зумовило як зростання їхньої міцності, так і забезпечило виконання ряду специфічних функцій. Один з можливих шляхів зміцнення міжклітинних контактів — збільшення площі контактуючих ділянок двох сусідніх клітин. При цьому пальцевидні вирости плазмолеми і цитоплазми однієї клітини занурюються у відповідні заглибини плазмолеми сусідньої клітини. Такий тип контакту носить назву пальцевидного, зубчастого, або контакту за типом замка. Відстань між плазмолемами контактуючих клітин при цьому така ж, як і в зоні простого контакту — 10...20 нм.
У процесі еволюційного розвитку багатоклітинних систем значно ускладнились форми і види міжклітинних контактів, що зумовило як зростання їхньої міцності, так і забезпечило виконання ряду специфічних функцій. Один з можливих шляхів зміцнення міжклітинних контактів — збільшення площі контактуючих ділянок двох сусідніх клітин. При цьому пальцевидні вирости плазмолеми і цитоплазми однієї клітини занурюються у відповідні заглибини плазмолеми сусідньої клітини. Такий тип контакту носить назву пальцевидного, зубчастого, або контакту за типом замка. Відстань між плазмолемами контактуючих клітин при цьому така ж, як і в зоні простого контакту — 10...20 нм.
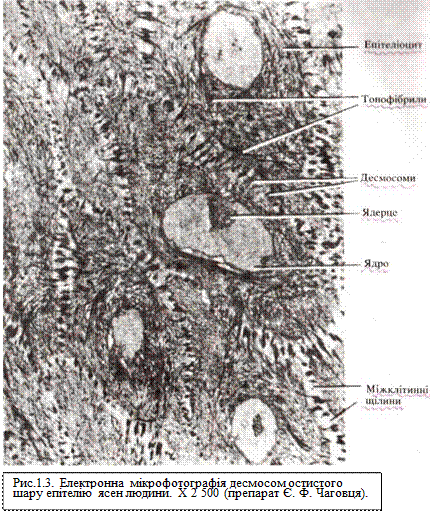
Подальше зміцнення зв'язку між клітинами досягається шляхом іммобілізації (знерухомлення) поверхні сусідніх ділянок плазмолеми контактуючих клітин (утворення так званих пластинок прикріплення) за допомогою фібрилярних органел цитоплазми і кортикального шару плазмолеми (рис. 1.3). Такий тип зв'язку між клітинами має назву десмосоми і зустрічається там, де необхідно досягти максимальної міцності міжклітинних зв'язків, наприклад, у складі епітеліальної тканини поверхні тіла. Міжклітинна щілина в ділянці десмосоми заповнена електронно-щільною речовиною, у якій розрізняють особливі фібрилярні структури, стабілізуючі даний тип контакту. У ділянці десмосомних контактів ширина міжклітинної щілини становить близько 25-30 нм, діаметр десмосоми — 0,5 мкм. У місцях контакту епітеліальних клітин з базальною мембраною утворюються структури, які мають назву напівдесмосом. Якщо десмосома складається з двох, то напівдесмосома — лише з однієї пластинки прикріплення.
Наступна форма контакту — з утворенням щільних замикаючих пластин, або щільний замикаючий контакт. У ділянці такого контакту відбувається максимальне зближення плазматичних мембран сусідніх клітин. Виступаючі кінці інтегральних білків плазмолем сусідніх клітин стикуються між собою, а існуючий проміжок ущільнюється за рахунок анастомозуючих фібрил та іонів кальцію. Зовнішні гідрофільні шари і глікокалікс суміжних плазмолем ніби зливаються при цьому в один суцільний шар завтовшки 2...3 нм. Щільний замикаючий контакт характерний для апікальної поверхні клітин, що вистеляють травний канал. Внаслідок утворення замикаючих пластин досягається повне відмежування міжклітинного простору від зовнішнього середовища. Щільні замикаючі контакти зустрічаються у всіх видах епітеліїв (ендотелій, мезотелій, епендима, кишковий епітелій). Контакти цього типу знайдені між фібробластами, ембріональними клітинами ектодерми і мезенхіми тощо.
Щілинний контакт, або нексус, забезпечує безпосередній обмін речовин між сусідніми клітинами. У зонах цих контактів, які мають розміри від 0,5 до 5 мкм, гексагонально розміщені частинки — конексони з діаметром 7...8 нм і каналом шириною близько 1,5 нм у центрі. Кожний конексон складається з шести субодииць білка конектину. Конексони вмонтовані в мембрану так, що пронизують її наскрізь. Співпадаючі на плазмолемах двох сусідніх клітин канали двох конексонів замикаються «кінець в кінець», внаслідок чого встановлюється безпосередній хімічний зв'язок між цитоплазмами сусідніх клітин: зв'язані щілин ними контактами клітини можуть вільно обмінюватися малими молекулами (неорганічними іонами, цукрами, амінокислотами, нуклеотидами, вітамінами), маса яких не перевищує 1000... 1500 дальтон. При цьому досягається своєрідна метаболічна кооперація клітин. У ділянках утворення щілинних контактів плазмолеми суміжних клітин зближені на відстань до 2...4 нм. Щілинними контактами зв'язані, зокрема, м'язові клітини міокарда, гладкі міоцити м'язової оболонки матки, овоцити і фолікулярні клітини яєчника тощо.
С и н а п с — спеціалізований контакт між нервовими клітинами або між нервовою клітиною і м'язом, у зоні якого відбувається передача нервового імпульсу. Основні структурні компоненти синапса: пресинаптична мембрана (ділянка плазмолеми відростка нервової клітини, з якої надходить сигнал), постсинаптична мембрана (ділянка плазмолеми клітини, яка сприймає сигнал), синаптична щілина шириною 20...30 нм (розмежовує пре- і постсинаптичну мембрани), заповнені нейромедіатором синаптичні пухирці. Функціонування синапсів забезпечує односторонню передачу інформації від клітини до клітини за допомогою медіатора (хімічного посередника).
Таким чином, виходячи з характеристик міжклітинних контактів, їх можна умовно поділити на три групи: адгезивні (зв'язуючі), ізолюючі та комунікаційні. До першої групи належать: простий адгезивний контакт, контакт за типом замка та десмосомний контакт. До другої групи належить щільний замикаючий контакт, до третьої — щілинний (нексус) та синаптичний контакти.
Дата добавления: 2014-12-03; просмотров: 681; Мы поможем в написании вашей работы!; Нарушение авторских прав |