КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
КРОВОТВОРЕННЯ (ГЕМОЦИТОПОЕЗ)
Формені елементи крові, які здебільшого є високоспеціалізованими клітинами, мають обмежений термін життя. Наприклад, еритроцити живуть близько 120 діб, гранулоцити перебувають у крові 10...20 год, а в тканинах 24...48 год, моноцити циркулюють у крові 30...60 год, тромбоцити живуть дві-три доби. Сталість якісного та кількісного складу формених елементів крові досягається їх постійним утворенням, розвитком, що й позначають терміном гемоцитопоез (від грецького «гайма» - кров, «цитос» - клітина, «поезіс» - творення), або кровотворення. У процесі кровотворення компенсується природна втрата віджилих формених елементів, тому гемопоез можна розглядати як процес фізіологічної регенерації крові.
Після народження кровотворення відбувається в органах, які мають назву кровотворних. До них належать червоний кістковий мозок плоских та епіфізів довгих трубчастих кісток — тут утворюються еритроцити, гранулоцити, моноцити, тромбоцити і попередники лімфоцитів; селезінка, лімфатичні вузли, тимус — у цих органах здійснюється диференціація і розмноження Т- і В-лімфоцитів і плазмоцитів. Гемопоетичну тканину червоного кісткового мозку називають мієлоїдною, а процес утворення еритроцитів, гранулоцитів, моноцитів і тромбоцитів позначають терміном мієлопоез. Кровотворну тканину, яка розташована в селезінці, лімфатичних вузлах і тимусі (а також систему цих органів), називають лімфоїдною, а процес утворення в них лімфоцитів і плазмоцитів — лімфопоезом.
Різні теорії кровотворення, які існували до недавніх часів, базуються на виділенні однієї або кількох родоначальних клітин, з яких утворюються усі види зрілих формених елементів. Поліфілетичні теорії, згідно з якими існують дві, три і більше вихідних клітинних форм (вони мали назви — дуалістична, триалістична та ін.) у даний час мають лише історичний інтерес. Тепер загальновизнаною є унітарна теорія кровотворення, згідно з якою всі зрілі формені елементи крові походять з одної загальної родоначальної клітини. Вперше основи цієї теорії сформулював ще на початку XX ст. російський гістолог О.О.Максимов, який вважав, що така клітина існує і має морфологію малого лімфоцита. У наш час ці уявлення підтверджені численними експериментами, які базуються на нових методах досліджень і дають змогу отримувати клітинні клони (група клітин, що утворюються з одної клітини), або кровотворні колонії, у селезінці смертельно опромінених мишей (метод колоніє утворення Тіл — Мак Кулох, 1961). Дані, отримані в цих дослідах, лягли в основу сучасної унітарної теорії кровотворення, згідно якої всі зрілі формені елементи крові походять з єдиної вихідної клітини, яку називають стовбуровою кровотворною клітиною (СКК).
Популяція СКК має такі ознаки: 1) поліпотентність, тобто здатність диференціюватися у напрямках усіх видів формених елементів крові; 2) здатність до самопідтримання протягом часу, близького до терміну існування самого організму людини: число мітозів, яке здіснює одна клітина, може перевищувати 100; 3) незважаючи на високу здатність до проліферації, стовбурова клітина у нормі поділяється дуже рідко, перебуваючи у G0-фазі клітинного циклу, однак під дією, наприклад, радіації вона може дуже скоро почати проліферацію; 4) СКК знаходяться у стані постійної й інтенсивної репопуляції, тобто мігрують з одних кровотворних органів у інші через кров; доказом цього є факт, що СКК завжди можна знайти у крові у вигляді клітин, здатних відновити гемопоез опромінених тварин.
У дорослих ссавців СКК скупчені головним чином у червоному кістковому мозку (на 105 ядерних клітин кісткового мозку припадає 50 СКК). Загальна кількість стовбурових кровотворних клітин у людини становить приблизно 5•1010 (третина з них знаходиться у мітотичному циклі). Виникають стовбурові клітини крові в ембріональному періоді у жовтковому мішку (знайдені вперше у семи-восьмиденних ембріонів мишей) і потім розселяються у всій кровотворній системі. СКК дорослих тварин є їхніми нащадками.
Морфологічно СКК не ідентифіковані, що пов'язано з їх малою концентрацією в кістковому мозку (10-3 – 10-4). Описано морфологію так званого кандидата у стовбурову клітину крові, отриманого спеціальними методами згущення. Ця клітина подібна до лімфоцитів кісткового мозку. Форма її кругла або овальна, діаметр 8 мкм, ядерно-цитоплазматичне відношення більше трьох (тобто клітина ядерного типу). Діаметр ядра 5 мкм, воно овальне або кругле з хвилястими краями, із щільними скупченнями хроматину біля ядерної мембрани. Обідок цитоплазми тонкий, багато поодиноких рибосом, невелика кількість мітохондрій та канальців гранулярної ендоплазматичної сітки; комплекс Гольджі спостерігається дуже рідко.
Згідно з сучасною схемою кровотворення у всіх гістогенетичних рядах, що завершуються утворенням зрілих формених елементів крові, виділяють такі класи клітин:
I клас — поліпотентні клітини-попередники (представлений СКК);
IIклас — частково детерміновані клітини-попередники (потенції цих клітин частково обмежені щодо дальшої їх диференціації, тобто з них можуть утворюватися вже не всі види формених елементів);
III клас — уніпотентні клітини-попередники (ці клітини здатні розвиватися лише в одному напрямку під впливом гормоноподібних речовин, які мають назву гемопоетинів;в різних гістогенетичних рядах існують різні гемопоетини);
IV клас — морфологічно розпізнавані проліферуючі клітини-попередники (на відміну від клітин перших трьох класів, які морфологічно не ідентифіковані, існування яких доведено лише експериментальним шляхом, клітини IV класу можна розпізнати на мазках кісткового мозку, вони здатні до мітотичного поділу);
V клас — дозріваючі клітини (втрачають здатність до мітотичного поділу і зазнають змін, пов'язаних з їх перетворенням у зрілі формені елементи);
VI клас — зрілі клітини, здатні до виходу у кров.
Окремі гістогенетичні ряди мають такі назви: еритропоез, гранулоцитопоез, моноцитопоез, тромбоцитопоез, лімфопоез.
Еритропоез (розвиток еритроцитів) відбувається у постнатальному періоді в червоному кістковому мозку. Джерело розвитку — стовбурова кровотворна клітина (І клас). Під впливом специфічного мікрооточення строми кісткового мозку ця клітина, поділяючись, диференціюється у клітину-попередника мієлопоезу (II клас, частково детермінована; з неї можуть утворитися тільки мієлоїдні елементи). Цю клітину ще позначають як колонієутворюючу одиницю гранулоцитів, еритроцитів, моноцитів, мегакаріоцитів (КУО-ГЕММ), або напівстовбурову клітину (НСК). З цієї клітини утворюються більш детерміновані попередники двох видів (це стосується еритропоезу): КУО-ГнЕ (колонієутворююча одиниця нейтрофільних гранулоцитів та еритроцитів) і КУО- МГЦЕ (колонієутворююча одиниця мегакаріоцитів та еритроцитів). Таким чином, наступна стадія розвитку еритроцитів — їх уніпотентний попередник КУО-Е (клітини III класу, розвиваються тільки в напрямку еритроцитів) може утворитися двома шляхами — з КУО-ГнЕ або КУО-МГЦЕ. Уніпотентну клітину еритропоезу називають ще еритропоетинчутливою (ЕЧК), тому що її подальша диференціація індукується гормоном еритропоетином. Останній виробляється у нирках і посилює проліферацію ЕЧК та їх перетворення в проеритробласти. Цей гормон також стимулює розвиток і розмноження еритроїдних клітин подальших стадій.
Проеритробласти (IV клас) (рис. 2.8) — перші морфологічно розпізнавані клітини еритроїдного ряду. Мають круглу форму, великі за розміром (діаметр клітин 15...25 мкм). Ядро велике, кругле, розташоване центрально, має дрібносітчасто-зернисту структуру, містить одно-три ядерця; цитоплазма забарвлюється базофільно, навколо ядра знаходиться світла перинуклеарна зона, є багато рибосом, невелика центросома з двома центріолями, характерною є наявність зерен феритину (комплекс білка з залізом) і виростів цитоплазми. Проеритробласти поділяються і перетворюються в базофільні еритробласти.
Базофільні еритробласти мають трохи менші розміри порівняно з проеритробластами (10...18 мкм). Хроматин у ядрі починає розташовуватися грудочками променеподібно, як спиці в колесі. Цитоплазма інтенсивно базофільна внаслідок великої кількості РНК.
У цих клітинах розпочинається синтез гемоглобіну. Вони поділяються мітозом і, нагромадивши певну кількість гемоглобіну, перетворюються в поліхроматофільні еритробласти.
Поліхроматофільні еритробласти менші за розмірами (10...14 мкм), ядро менше, більш щільне, з чіткою колесоподібною структурою хроматину. Ядерця не 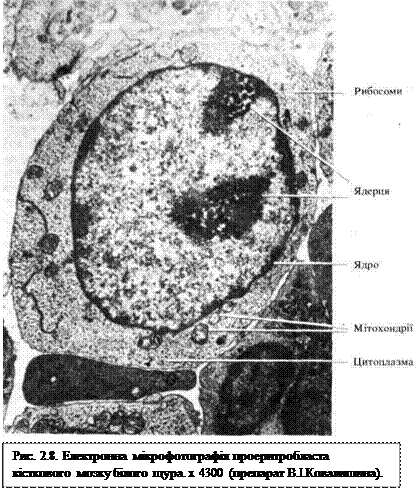 визначаються. Цитоплазма забарвлюється поліхромно, тобто і кислими і основними барвниками. Оксифілія зумовлена наявністю гемоглобіну, а базофілія — наявністю РНК. Гемоглобін у цих клітинах може розташовуватися дифузно, тоді вся цитоплазма забарвлюється у сіруватий колір або плямами, які сприймають кислі барвники, або у вигляді обідка навколо ядра. Число рибосом у цих клітинах зменшується, феритин розміщується агрегатами. Поліхроматофільні еритробласти поділяються мітозом. Їх пізні генерації називаються поліхроматофільними нормобластами.
визначаються. Цитоплазма забарвлюється поліхромно, тобто і кислими і основними барвниками. Оксифілія зумовлена наявністю гемоглобіну, а базофілія — наявністю РНК. Гемоглобін у цих клітинах може розташовуватися дифузно, тоді вся цитоплазма забарвлюється у сіруватий колір або плямами, які сприймають кислі барвники, або у вигляді обідка навколо ядра. Число рибосом у цих клітинах зменшується, феритин розміщується агрегатами. Поліхроматофільні еритробласти поділяються мітозом. Їх пізні генерації називаються поліхроматофільними нормобластами.
Поліхроматофільні нормобласти мають розміри до 10 мкм, тобто менші, ніж попередники, ядро втрачає колесоподібне розташування хроматину і ущільнюється. Воно стає пікнотичним, майже безструктурним. Клітини втрачають здатність до поділу (V клас). 80% клітин на цій стадії втрачає ядро і перетворюється на кістково-мозкові ретикулоцити. Останні продовжують накопичувати гемоглобін. їхнє дозрівання у кістковому мозку продовжується протягом 36...44 год, потім вони надходять у кров у вигляді зрілих еритроцитів (VI клас). Частина кістково-мозкових ретикулоцитів залишає кістковий мозок не повністю насиченими гемоглобіном — це і є ретикулоцити крові. 20 % поліхроматофільних нормобластів, не втрачаючи ядра, продовжують накопичувати гемоглобін і перетворюються в оксифільні нормобласти. Їхня цитоплазма оксифільна. Вони втрачають ядро шляхом виштовхування його з клітини або відриву від клітини ядерного фрагмента (таким же шляхом втрачають ядро поліхроматофільні нормобласти) і перетворюються в еритроцити.
Весь процес утворення еритроцитів у здорових людей від проеритробласта до еритроцита продовжується шість-вісім днів. Диференціація ядерних елементів еритропоезу триває 100…140 год. Морфологічно розпізнавані клітини еритроїдного ряду здійснюють 5-6 мітозі в, а у попередніх класах — 10-15 мітозів. З кожного проеритробласта утворюється 30-60 еритроцитів. Загальне число еритроїдних клітин у кістковому мозку людини дорівнює З•1011.
Таким чином, у процесі розвитку еритроцитів від проеритробласта до зрілої клітини проходять такі основні зміни: 1) базофілія цитоплазми, зумовлена наявністю значної кількості РНК у рибосомах, змінюється на поліхроматофілію, а потім на оксифілію внаслідок збільшення кількості гемоглобіну і зменшення кількості РНК; втрачаються усі органели; 2) ядро ущільнюється, пікнотизується і виштовхується; 3) розміри клітини у процесі диференціації зменшуються від 15...25 до 7...8 мкм.
У нормі потреба в еритроцитах забезпечується за рахунок посиленого розмноження поліхроматофільних еритробластів. Якщо потреба організму в еритроцитах зростає (наприклад, при крововтратах), еритробласти починають розвиватися із попередників, а останні — зі стовбурових клітин.
Гранулоцитопоез (розвиток гранулоцитів).Першими клітинами гранулоцитопоезу є стовбурова кровотворна клітина червоного кісткового мозку і клітина-попередник мієлопоезу, які аналогічні вищеописаним для розвитку еритроцитів (І і IIкласи). Наступним етапом є утворення більш детермінованої клітини-попередника гранулоцитів і моноцитів-макрофагів, або КУО-ГМ. З неї при розвитку гранулоцитів утворюються уніпотентні попередники (III клас): базофілів (КУО-Б), еозинофілів (КУО-Ео) та нейтрофілів (КУО-Гн). Крім того, уніпотентний попередник нейтрофілів може утворюватися також із КУО-ГнЕ (колоніє-утворююча одиниця нейтрофілів та еритроцитів). Гормон, який стимулює диференціацію та проліферацію клітин даного ряду, має назву гранулопоетину.
Першою морфологічно розпізнаваною клітиною цього ряду є мієлобласт (IV клас). Вона велика (до 20 мкм), кругле ядро розташоване у центрі, займає більшу частину клітини, цитоплазма її базофільна. Ядро має ніжносітчасту структуру (на відміну від еритробласту не містить зерен хроматину), має від двох до п'яти ядерець синього кольору. У цитоплазмі багато рибосом, мітохондрій, можна виявити неспецифічну азурофільну зернистість. Експериментально доведено, що морфологічно ідентичні мієлобласти уже комітовані тільки до одного шляху диференціації і серед них є базофільні, еозинофільні та нейтрофільні мієлобласти. Вони поділяються мітозом один раз і перетворюються у промієлоцити.
Промієлоцити мають розміри 12...20 мкм, на відміну від мієлобластів мають згрубілу структуру ядра, меншу кількість неспецифічної зернистості. Утворення промієлоцитів супроводжується появою специфічної зернистості, залежно від її характеру, розрізняють три типи цих клітин — базофільні, еозинофільні та нейтрофільні. Промієлоцити здійснюють один мітоз і перетворюються у мієлоцити.
Мієлоцити мають розміри 8...12 мкм. Ядро містить щільні хроматинові тяжі, які чергуються зі світлими ділянками, ядерця відсутні. Цитоплазма слабо базофільна або слабо оксифільна (тобто базофілія цитоплазми зменшується), наростає число специфічних зерен. Серед мієлоцитів чітко визначаються три різновиди: базофільні, еозинофільні та нейтрофільні. Ці клітини поділяються мітозом двічі, співвідношення ранніх і пізніх мієлоцитів становить 1:2. Перетворюються в метамієлоцити.
Метамієлоцити — круглі клітини діаметром близько 8 мкм, об'єм цитоплазми переважає над об'ємом ядра. Ядро бобовидної або підковоподібної форми. Ці клітини вже не поділяються і тому належать до класу дозріваючих клітин (V клас). Існує три види цих клітин — базофільні, еозинофільні та нейтрофільні. Метамієлоцити можуть попадати у периферійну кров і тоді називаються юними.
Із метамієлоцитів утворюються паличкоядерні гранулоцити шляхом зміни форм ядра (воно видовжується і вигинається). У зрілих сегментоядерних гранулоцитів ядро поділяється на сегменти (VI клас). Найбільше сегментується ядро у нейтрофільних гранулоцитів і найменше — у базофілів.
Таким чином, при утворенні гранулоцитів у клітинах відбуваються такі морфологічні зміни: зменшення розмірів клітини; зменшення базофілії цитоплазми; порушення ядерно-цитоплазматичного співвідношення у бік зростання кількості цитоплазми; ущільнення і зміна форми ядра; накопичення специфічної зернистості.
Моноцитопоез (розвиток моноцитів). Клітини-попередники моноцитів перших двох класів були описані вище. З клітини-попередника гранулоцитів і моноцитів-макрофагів утворюється уніпотентний попередник моноцитів, або КУО-М (III клас). Перша морфологічно розпізнавана клітина — моноцитобласт (IV клас). Це велика клітина (до 22 мкм) з круглим ядром і вузькою облямівкою базофільної цитоплазми. Поділяючись, вона диференціюється в промоноцит, а останній перетворюється у моноцит. При цьому з клітиною відбуваються такі зміни: збільшується кількість цитоплазми, її базофілія дещо зменшується, а ядро набуває бобовидної форми. Ядерно-цитоплазматичне співвідношення у моноцита дорівнює 1:1. Моноцити, однак, не є кінцевою стадією диференціації цього ряду і перетворюються далі у макрофаги (гістіоцити-макрофаги) сполучної тканини. На шляху від моноцитобласта до макрофага відбувається сім-вісім мітозів.
Тромбоцитопоез (розвиток кров'яних пластинок). Перші два класи клітин-попередників тромбоцитопоезу описані вище при викладі еритропоезу (СКК – КУО – ГЕММ – КУО – МГЦЕ). Уніпотентний попередник — це колонієутворююча одиниця мегакаріоцит і в (КУО-МГЦ), або тромбоцитопоетинчутлива клітина за назвою фактора гормонального типу, який діє у цьому гістогенетичному ряді.
 Мегакаріобласт — наймолодша морфологічно розпізнавана клітина тромбоцитопоезу. Вона кругла, розміри її 25...40 мкм. Ядро з рівномірним розподілом хроматину насиченого фіолетового кольору містить одне — три ядерця. Цитоплазма базофільна, темно-синього кольору, має дві зони: перину-клеарну, яка містить органели, і периферійну, пронизану вгинаннями плазмолеми, що утворюють складні демаркаційні трубочки. З мегакаріобласту виникає промегакаріоцит. Розміри його 40...80 мкм. Ядро часто бухтоподібне, починається його сегментація і огрубіння структури. Цитоплазма стає менш базофільною, з'являється азурофільна зернистість, трубочки демаркаційної системи виникають не тільки на периферії, а й у середній зоні цитоплазми. Цромегакаріоцит диференціюється далі у мегакаріоцит.
Мегакаріобласт — наймолодша морфологічно розпізнавана клітина тромбоцитопоезу. Вона кругла, розміри її 25...40 мкм. Ядро з рівномірним розподілом хроматину насиченого фіолетового кольору містить одне — три ядерця. Цитоплазма базофільна, темно-синього кольору, має дві зони: перину-клеарну, яка містить органели, і периферійну, пронизану вгинаннями плазмолеми, що утворюють складні демаркаційні трубочки. З мегакаріобласту виникає промегакаріоцит. Розміри його 40...80 мкм. Ядро часто бухтоподібне, починається його сегментація і огрубіння структури. Цитоплазма стає менш базофільною, з'являється азурофільна зернистість, трубочки демаркаційної системи виникають не тільки на периферії, а й у середній зоні цитоплазми. Цромегакаріоцит диференціюється далі у мегакаріоцит.
Мегакаріоцит (рис. 2.9) — найбільша клітина червоного кісткового мозку. її розміри від 50...70 до 100 мкм. Ядро поліморфне, фрагментоване, із заглибленнями і вирізами, дуже химерної (примхливої) форми, структура його грубосітчаста, ядерець немає. Цитоплазма базофільна, забарвлюється у фіолетовий або рожево-фіолетовий колір, містить азурофільну зернистість. Клітина нечітко відмежована від оточуючого середовища. За ходом демаркаційних трубочок цитоплазма розбивається на невеличкі фрагменти, які відокремлюються від клітини, перетворюючись на кров'яні пластинки (з одної клітини утворюється три-чотири тисячі тромбоцитів). Особливість мегакаріоцитів полягає також у тому, що ці клітини є поліплоїдними, причому число хромосомних наборів у них може досягати 32-64.
Поліплоїдизація цих клітин зумовлена тим, що на шляху їхнього утворення з мегакаріобластів поділи не відбуваються, а здійснюються чотири-п'ять ендомітозів, у результаті чого збільшується об'єм і ядра, і цитоплазми.
Мегакаріоцити локалізуються екстраваскулярно, прилягаючи до ендотеліальних клітин, а їхні цитоплазматичні відростки проникають у просвіт синуса. Відростки бувають двох типів. Одні не містять органел і фіксують мегакаріоцит до ендотелію. Інші мають розміри 2,5х120 мкм і, проникаючи у просвіт синусів, дають початок приблизно тисячі тромбоцитів. Можливо також, що мегакаріоцити регулюють міграцію інших кровотворних клітин через стінку синусів.
Лімфопоез (розвиток лімфоцитів). Згідно з унітарною теорією кровотворення джерелом розвитку лімфоцитів є стовбурова кровотворна клітина (І клас), з якої утворюються клітина-попередник лімфопоезу (IIклас). Далі розвиток цієї клітини іде у двох напрямках мідповідно до двох різновидів лімфоцитів — Т і В. В обох рядах виникають уніпотентні попередники, які через лімфобласти (Т і В) перетворюються у лімфоцити (Т і В).
Розвиток Т-лімфоцитів проходить у тимусі під впливом специфічного мікрооточення його строми і гормону цього органа. Розвиток В-лімфоцитів у людини здійснюється у червоному кістковому мозку і, можливо, у лімфатичних фолікулах травної трубки. Попередники Т- і В-лімфоцитів утворюються також у червоному кістковому мозку.
Особливістю цих рядів є те, що зрілі клітини не є кінцевими елементами і їхній подальший гістогенез залежить від присутності антигенів. Тоді вони переходять у бластні форми і починають поділ. При повторній антигенній стимуляції В-лімфоцити, наприклад, можуть давати клони з астрономічним числом клітин, здійснюючи до 90 мітозів. Цей, так званий антигензалежний процес диференціації лімфоцитів відбувається у периферійних кровотворних органах — селезінці та лімфатичних вузлах. Тут із стимульованих антигеном Т-лімфоцитів через Т-лімфобласти, великі та середні лімфоцити утворюються Т-кілери, Т-супресори, Т-клітини пам'яті. Стимульовані В-лімфоцити через плазмобласти і проплазмоцити трансформуються в плазмоцити (рис. 2.10) і В-клітини пам'яті.
Ембріональний гемопоез.У процесі ембріонального гемопоезу відбувається розвиток крові як тканини. У людини кровотворення вперше спостерігається на другому-третьому тижні ембріонального розвитку в стінці жовткового мішка. Спочатку тут виникають ущільнені ділянки мезенхіми — кров'яні острівці. Клітини на периферії острівця стають плоскими, сполучаються між собою і утворюють судинну стінку. Центральні клітини втрачають відростки, заокруглюються і перетворюються у СКК. Частина СКК диференціюється у первинні клітини крові (бласти) — великі клітини з базофільною цитоплазмою і великими, добре помітними ядерцями в ядрі. Ці клітини мітотично поділяються і перетворюються в первинні еритробласти (мегалобласти) — великі ядерні клітини з базофільною цитоплазмою. Вони швидко накопичують гемоглобін, перетворюючись на оксифільні еритробласти, а останні втрачають ядро і стають первинними еритроцитами — мегалоцитами. Але втрата ядра відбувається не у всіх клітин: частина первинних еритроцитів функціонує у вигляді ядерних клітин. Таким чином, еритроцити на цьому етапі ембріогенезу утворюються скороченим шляхом, мають великі розміри, виникають всередині судин (інтраваскулярно). Такий тип кровотворення має назву мегалобластичного і є нормою для ембріогенезу. Поява такого кровотворення у постнатальному періоді свідчить про патологію (злоякісна анемія). Поряд з мегалобластичним (у стінці жовткового мішка) починається процес нормобластичного кровотворення, який веде до появи еритроцитів-нормоцитів. Крім того, тут екстраваскулярно з частини первинних клітин-бластів утворюється невелика кількість гранулоцитів — нейтрофілів та еозинофілів. Описаний тип ембріонального кровотворення отримав назву мезобластичного (позазародкового).
Частина СКК лишається у недиференційованому стані, розноситься з кров'ю до різних органів зародка, в яких починається процес кровотворення. Наприклад, на п'ятому тижні ембріогенезу кровотворення починається у печінці. Тут утворюються еритроцити і гранулоцити, головним чином нейтрофільні та еозинофільні. Розвиток іде екстраваскулярно. Крім гранулоцитів, у печінці формуються гігантські' клітини —  мегакаріоцити. Процес кровотворення у печінці припиняється до кінця внутріутробного періоду.
мегакаріоцити. Процес кровотворення у печінці припиняється до кінця внутріутробного періоду.
На початку другого місяця ембріогенезу виникає тимус, а на сьомому-восьмому тижні він заселяється стовбуровими клітинами, з яких утворюються перші лімфоцити. На третьому місяці кровотворення починається у селезінці. Тут із стовбурових клітин утворюються всі формені елементи крові. Таким чином, селезінка являє собою в ембріогенезі універсальний кровотворний орган. Після п'ятого місяця в ній починає переважати лімфопоез. Цей період ембріонального гемопоезу отримав назву гепато-тимолієнального.З четвертого місяця ембріогенезу починає функціонувати кістковий мозок, а з шостого місяця він стає основним універсальним органом кровотворення. У цьому періоді гемопоез іде також у тимусі, лімфатичних вузлах і селезінці, внаслідок чого його називають медуллотимо-лімфоїдним.
Дата добавления: 2014-12-03; просмотров: 1036; Мы поможем в написании вашей работы!; Нарушение авторских прав |