КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
СПОЛУЧНІ ТКАНИНИ ЗІ СПЕЦІАЛЬНИМИ ВЛАСТИВОСТЯМИ
Сполучна тканина (textus connectivus) — дуже поширена в організмі: загалом вона становить близько 50 % маси тіла. Зі сполучної тканини побудовані скелет, шкіра, хрящі, сухожилля та зв'язки, основа органів. Сполучну тканину поділяють на власне сполучну, хрящову і кісткову. Власне сполучна тканина, у свою чергу, поділяється на волокнисту і сполучні тканини зі спеціальними властивостями. До останніх належить ретикулярна, жирова, пігментна та слизова тканини. Волокниста, сполучна тканина залежно від вмісту волокнистих структур є пухкою і щільною. Пухка містить порівняно більше клітин і аморфної речовини, а щільна багатша на волокнисті структури. Щільну сполучну тканину залежно від розташування волокнистих структур поділяють на оформлену та неоформлену: в оформленій волокна розташовані паралельно, а в неоформленій ідуть у різних напрямках, утворюючи сітку.
Серед усіх згаданих у класифікації різновидів сполучної тканини найбільш поширеною й такою, що містить усі види елементів, є пухка волокниста сполучна тканина. Вона міститься майже у всіх внутрішніх органах, утворює їхні оболонки, заповнює проміжки між органами, підстилає епітелій, супроводжує судини та нерви. Вона виконує всі функції, які властиві тканинам внутрішнього середовища, а саме: трофічну, захисну, опорно-механічну. Крім того, пухка сполучна тканина виконує також замісну функцію (при пошкодженні заміщає, заповнює собою дефект в органах).
Пухка сполучна тканина (textus connectivus laxus) побудована з клітин і міжклітинної речовини (рис. 2.11). Остання, у свою чергу, включає волокнисті структури (колагенові, еластичні і ретикулярні волокна) й основну міжклітинну речовину. Подібний план будови характерний і для всіх інших різновидів сполучної тканини. До клітинних елементів пухкої сполучної тканини належать: фібробласти, макрофаги, плазмоцити, тканинні базофіли, адипоцити, піг-ментоцити, адвентиційні клітини, а також лейкоцити, які мігрують з крові.
Клітинні елементи сполучної тканини. Фібробласти — це клітини-продуценти міжклітинної речовини. Саме вони синтезують як волокнисті структури, так і основні компоненти аморфної речовини. У певному розумінні фібробласти будують сполучну тканину. За їхньою властивістю будувати основні опорні структури організму фібробласти часто називають механоцитами. Про здатність створювати волокна свідчить їхня назва («фібра» — волокно та «бластос» — зачаток). Діяльністю цих клітин зумовлене загоювання ран, розвиток рубця, утворення капсули навколо стороннього тіла тощо. До фібробластів належить численна група клітин, різних за ступенем диференціації, які утворюють так званий фібробластичний ряд (або диферон): стовбурові клітини — напівстовбурові клітини-попередники-мало-спеціалізовані фібробласти-зрілі фібробласти-фіброцити. Крім того, до цього ж ряду належать міофібробласти та фіброкласти.
Малоспеціалізовані, або юні, фібробласти округлої або веретеноподібної форми з базофільною цитоплазмою містять велику кількість вільних рибосом. Інші органели (ендоплазматична сітка, мітохондрії, комплекс Гольджі) розвинені слабо, здатні до мітотичного поділу. Мають низький рівень синтезу і секреції білка. Розміри їх не перевищують 20...25 мкм.
 Зрілі (спеціалізовані) фібробласти (рис. 2.12) — великі клітини з відростками. На препараті-плівці у розпластаному вигляді вони можуть досягати 40...50 мкм і більше, товщина їх незначна. Вони плоскі, пластинчасті. Ядро цих клітин велике, овальне, світле, містить дрібнорозпилений рівномірно розподілений хроматин, на тлі якого добре видно одне-два великих ядерця. Цитоплазма забарвлюється базофільно. На плівковому препараті можна бачити розподіл клітинного тіла фібробласта на дві зони — внутрішню ендоплазму, яка забарвлюється інтенсивніше, і зовнішню ектоплазму, забарвлення якої значно слабше; вона не має чітких меж і зливається з оточуючою міжклітинною речовиною. Цитоплазма фібробласта містить усі загальні органели. Особливо добре розвинена гранулярна ендоплазматична сітка, яка займає до 35 % об'єму клітини; тут відбувається синтез проколагену, еластину. Добре розвинений також і комплекс Гольджі, який становить близько 10 % об'єму клітини, має вигляд цистерн і пухирців, розкиданих по всій клітині; тут синтезуються глікозаміноглікани. Останні, як і фібрилярні білки, секретуються у міжклітинну речовину і входять до складу волокон і аморфної речовини. Фібробласти також синтезують фібрилярний глікопротеїн позаклітинного матриксу — фібронектин, який забезпечує зв’язування клітин з їхнім мікрооточенням і регулює пересування клітин. Мітохондрії великі, кількість їх помірна, як і лізосом.
Зрілі (спеціалізовані) фібробласти (рис. 2.12) — великі клітини з відростками. На препараті-плівці у розпластаному вигляді вони можуть досягати 40...50 мкм і більше, товщина їх незначна. Вони плоскі, пластинчасті. Ядро цих клітин велике, овальне, світле, містить дрібнорозпилений рівномірно розподілений хроматин, на тлі якого добре видно одне-два великих ядерця. Цитоплазма забарвлюється базофільно. На плівковому препараті можна бачити розподіл клітинного тіла фібробласта на дві зони — внутрішню ендоплазму, яка забарвлюється інтенсивніше, і зовнішню ектоплазму, забарвлення якої значно слабше; вона не має чітких меж і зливається з оточуючою міжклітинною речовиною. Цитоплазма фібробласта містить усі загальні органели. Особливо добре розвинена гранулярна ендоплазматична сітка, яка займає до 35 % об'єму клітини; тут відбувається синтез проколагену, еластину. Добре розвинений також і комплекс Гольджі, який становить близько 10 % об'єму клітини, має вигляд цистерн і пухирців, розкиданих по всій клітині; тут синтезуються глікозаміноглікани. Останні, як і фібрилярні білки, секретуються у міжклітинну речовину і входять до складу волокон і аморфної речовини. Фібробласти також синтезують фібрилярний глікопротеїн позаклітинного матриксу — фібронектин, який забезпечує зв’язування клітин з їхнім мікрооточенням і регулює пересування клітин. Мітохондрії великі, кількість їх помірна, як і лізосом.
У периферійному шарі цитоплазми розташовані мікрофіла-менти товщиною 5...6 нм, які містять скоротливі білки типу актину і міозину та зумовлюють здатність цих клітин до руху. Вважають, що серед фібробластів існують дві популяції: з коротким життєвим циклом (кілька тижнів) і з довгим життєвим циклом (кілька місяців).
Фіброцити — це дефінітивні (кінцеві) форми розвитку фібробластів. Форма їх веретеноподібна, вони можуть мати крилоподібні відростки. Містять невелику кількість органел. Синтетичні процеси в них різко знижені.
Міофібробласти — це вид клітин, у які можуть перетворюватися фібробласти. Вони функціонально подібні до гладких м’язових клітин, але, на відміну від останніх, мають добре розвинену ендоплазматичну сітку. Такі клітини можна спостерігати у матці під час вагітності, а також у грануляційній тканині (при загоюванні ран).
Фіброкласти — ще один вид клітин, похідних фібробластів. Вони мають високу фагоцитарну активність, містять значну кількість лізосом. Беруть участь у лізисі міжклітинної речовини; їх можна спостерігати у матці після закінчення вагітності.
Макрофаги (макрофагоцити).Ці клітини також називають гістіоцитами, клазматоцитами. За кількісним вмістом у пухкій сполучній тканині макрофаги займають друге місце після фібробластів. Порівняно з останніми вони мають менші розміри клітинного тіла (10...15 мкм), яке добре відмежоване від основної речовини. Форма різна: округла, витягнута або неправильна. Ядро теж має менші розміри, не таку правильну форму, як у фібробласта, містить більше гетерохроматину, виглядає щільним, забарвлюється досить інтенсивно. Цитоплазма макрофагів базофільна, неоднорідна, плямиста, містить багато лізосом, фагосом, піноцитозних пухирців. Інші органели (мітохондрії, гранулярна ендоплазматична сітка, комплекс Гольджі) розвинені помірно.
Цитоплазма макрофагів утворює глибокі складки і довгі мікроворсинки, за допомогою яких ці клітини захоплюють сторонні частинки. На поверхні плазмолеми макрофага містяться рецептори для пухлинних клітин, еритроцитів, Т- і В-лімфоцитів, антигенів, імуноглобулінів. Наявність рецепторів до імуноглобулінів забезпечує їхню участь в імунних реакціях.
Макрофаги відіграють важливу роль як у природному, так і в набутому імунітеті організму. Участь макрофагів у природному імунітеті виявляється у їхній здатності до фагоцитозу і в синтезі низки активних речовин — фагоцитину, лізоциму, інтерферону, пірогену, компонентів системи комплементу та інших, які є основними факторами природного імунітету. їхня роль у набутому імунітеті полягає в передачі антигену імунокомпетент-ним клітинам (лімфоцитам) після його переробки з корпускулярної форми в молекулярну (участь у кооперативній триклітинній системі імунної відповіді разом з Т- і В-лімфоцитами). Крім того, макрофаги секретують медіатори-монокіни, які сприяють специфічній реакції на антигени, і цитолітичні фактори, які вибірково руйнують пухлинні клітини.
Походять макрофаги з промо-ноцитів червоного кісткового мозку, тобто зі стовбурової гемопоетичної клітини, і завершують собою моноцитарний гістогенетичний ряд. Разом з інши  ми клітинами цього ж походження вони утворюють так звану макрофагічну систему організму.
ми клітинами цього ж походження вони утворюють так звану макрофагічну систему організму.
До макрофагічної системи належить сукупність усіх клітин, які здатні захоплювати з тканинної рідини організму сторонні частинки, загиблі клітини та неклітинні структури, бактерії тощо. Фагоцитований матеріал всередині клітини піддається ферментативному розщепленню в лізосомному апараті. Таким чином, ліквідуються шкідливі для організму агенти, які виникають місцево чи попадають іззовні. Ці клітини можна ідентифікувати за допомогою методу вітального забарвлення, використовуючи прижиттєве введення в організм трипанового синього, колоїдного срібла або китайської туші. Всі вказані колоїдні речовини фагоцитуються макрофагами завдяки тому, що утворюють макромолекуляр-ні агрегати, а клітини стають добре помітними на препараті. До таких клітин належать гістіоцити-макрофаги пухкої сполучної тканини, вільні та фіксовані макрофаги кровотворних органів, зірчасті клітини синусоїдних судин печінки, альвеолярні макрофаги легень, перитонеальні макрофаги, гліальні макрофаги нервової тканини (мікроглія), остеокласти, гігантські клітини сторонніх тіл. Усі вони здатні до активного фагоцитозу, мають на поверхні рецептори до імуноглобулінів (завдяки чому здатні до імунного фагоцитозу), походять з промоноцитів червоного кісткового мозку і моноцитів крові. На відміну від макрофагів, які І. І. Мечніков назвав «професійними фагоцитами», здатність до факультативного фагоцитозу мають інші види клітин — фібробласти, ретикулярні клітини, ендотеліоцити, нейтрофільні лейкоцити. Але ці клітини не належать до макрофагічної системи, тому що вони не можуть здійснювати специфічного імунного фагоцитозу, а також відрізняються своїм походженням.
Концепція фагоцитозу була уперше висунута І.І.Мечніковим. Він дійшов висновку, що фагоцитоз, який виник в еволюції як внутрішньоклітинне травлення і закріпився за багатьма клітинами, одночасно є важливим захисним механізмом. Він обгрунтував доцільність об'єднання таких клітин в одну систему і запропонував назвати її макрофагічною. У 30-50-х рр. цю захисну систему називали ретикулоендотеліальною (РЕС), помилково зараховуючи до неї деякі види факультативних фагоцитів. Останнім часом її називають системою мононуклеарних фагоцитів, що, однак, не зовсім точно, оскільки серед клітин цієї системи є і багатоядерні (остеокласти і гігантські клітини сторонніх тіл).
Макрофагічна система — міцний захисний апарат, який бере участь як у загальних, так і місцевих захисних реакціях організму. У цілісному організмі макрофагічна система регулюється місцевими механізмами, а також нервовою та ендокринною системами.
Плазматичні клітини (плазмоцити)(рис. 2.13) мають розміри 7...10 мкм, хоча можуть бути дещо більшими. Форма їх округла або багатокутна, якщо вони тиснуть одна на одну. Ядро невелике, кругле, розташоване ексцентрично, містить головним чином конденсований хроматин, грудочки якого утворюють характерний для плазмоцита малюнок — колеса зі спицями або цифр на циферблаті годинника. Цитоплазма інтенсивно баюфільна, на фоні якої біля ядра добре видно «світле подвір'я», або перинуклеарну зону із слабшим забарвленням. Ультраструктура цих клітин характеризується наявністю в цитоплазмі добре розвиненої гранулярної ендоплазматичної сітки, що розташована концентрично і займає більшу частину клітини. Велика кількість рибосом (РНК)зумовлює базофілію цитоплазми. У ділянці «світлого подвір'я» локалізовані центріолі, оточені цистернами комплексу Гольджі. У цистернах гранулярної ендоплазматичної сітки плазмоцитів відбувається синтез імуноглобулінів (антитіл). Частина вуглеводногокомпонента імуноглобулінів синтезується у комплексі Гольджі. Ця органела, яка досить добре розвинена у плазмоцитах, відповідає також за секрецію синтезованих імуноглобулінів за межі клітини: далі вони попадають через лімфу в кров.
Таким чином, плазмоцити забезпечують гуморальний імунітет тобто вироблення специфічних білків-антитіл, реагуючи на проникнення в організм антигену, який ними буде знешкоджуватись. Походять плазматичні клітини зі стовбурової кровотворної клітини, а саме з В-лімфоцитів. Плазматичні клітини здебільшого зустрічаються у пухкій сполучній тканині власного шару слизової оболонки кишки та дихальних шляхів, у лімфатичних вузлах, селезінці, в інтер-стиційній сполучній тканині різних залоз.
 Тканинні базофілимали багато назв, які доцільно навести, щоб допомогти орієнтуватися у літературі: мастоцити, лаброцити, тучні клітини. Останню назву дав цим клітинам П. Ерліх, який у 1877 р. вперше описав клітини, що були переповнені гранулами, ніби «об'їлися» ними. Ця назва дуже поширена у літературі. Назва «тканинні базофіли» відповідає сучасній міжнародній гістологічній номенклатурі й свідчить про те, що клітини мають зернистість, подібну до гранул базофільних лейкоцитів крові. Тканинні базофіли часто локалізуються за ходом кровоносних судин мікроциркуляторного русла, утворюючи периваскулярні піхви. Велика кількість цих клітин зустрічається у стінці органів шлунково-кишкового тракту, в матці, молочній залозі, тимусі, мигдаликах.
Тканинні базофілимали багато назв, які доцільно навести, щоб допомогти орієнтуватися у літературі: мастоцити, лаброцити, тучні клітини. Останню назву дав цим клітинам П. Ерліх, який у 1877 р. вперше описав клітини, що були переповнені гранулами, ніби «об'їлися» ними. Ця назва дуже поширена у літературі. Назва «тканинні базофіли» відповідає сучасній міжнародній гістологічній номенклатурі й свідчить про те, що клітини мають зернистість, подібну до гранул базофільних лейкоцитів крові. Тканинні базофіли часто локалізуються за ходом кровоносних судин мікроциркуляторного русла, утворюючи периваскулярні піхви. Велика кількість цих клітин зустрічається у стінці органів шлунково-кишкового тракту, в матці, молочній залозі, тимусі, мигдаликах.
Форма тканинних базофілів різноманітна так само, як і розміри. Вони бувають круглі, овальні, з широкими відростками. Розміри коливаються від 10...20 до 35 і навіть до 100 мкм. Ядра порівняно невеликі, круглі, звичайної будови. У цитоплазмі міститься велика кількість мітохондрій, небагато гранулярної, а також агранулярної ендоплазматичної сітки; комплекс Гольджі розвинений добре. Головна особливість цих клітин — наявність великої кількості характерних гранул розмірами 0,2...0,8 мкм, кожна з яких оточена мембраною. За електронно-мікроскопічною будовою гранули тканинних базофілів людини кристалоїдні або пластинчасті (спостерігаються видові відмінності структури гранул). Забарвлюється зернистість базофільно, метахроматично. Гранули містять кілька речовин, що мають велике фізіологічне значення. Першою з таких речовин є гепарин, який становить 30 % вмісту гранул і, головним чином, зумовлює їх базофілію і метахромазію. Друга речовина — гістамін, який становить 10 % їх вмісту. Матрикс гранули складається з білка (хімаза тучних клітин) та гепарину, які формують стабільну сітку; до неї іонними зв'язками приєднаний гістамін. Гранули також містять хондроїтинсульфат, гіалуронову кислоту, у деяких тварин знайдено також серотонін, але у людини його немає.
Гепарин — це сульфатований глікозаміноглікан, який уперше було виділено з печінки (цим зумовлена його назва) і який запобігає зсіданню крові. Виявлено, що тканинні базофіли синтезують гепарин у комплексі Гольджі. Вони можуть втрачати свої гранули (процес дегрануляції), і тоді гепарин виділяється у міжклітинну речовину. Гепарин знижує її проникливість, має протизапальну дію, є антикоагулянтом. Крім того, гепарин стимулює активність ферменту ліпопротеїнліпази і, таким чином, сприяє розпаду хіломікронів плазми.
Гістамін синтезується у тканинних базофілах за участю гістидиндекарбоксилази (маркерний фермент цих клітин), яка здійснює перетворення гістидину в гістамін, що діє на гладкі м'язи, викликаючи їхнє скорочення, а також сприяє виходу плазми з венул і капілярів за рахунок розширення і підвищення проникливості їх стінки. Внаслідок виходу плазми у пухкій сполучній тканині під епідермісом утворюються пухирі. Цей симптом отримав назву кропивниці. Описану дію гістаміну можна спостерігати під час анафілактичного шоку або алергії. Розвиток цих процесів і участь у них тканинних базофілів пояснюється так. У відповідь на проникнення в організм деяких антигенів, що звуться алергенами, утворюються специфічні антитіла, які належать до класу IgE. Тканинні базофіли, як і базофільні лейкоцити, мають рецептори для антитіл цього типу і зв'язують їх так, що варіабельні ділянки для зв'язування антигенів лишаються вільними. При повторному введенні антигену він швидко з'єднується зі специфічними антитілами на поверхні тучних клітин. Після утворення комплексу антиген-антитіло гістамін вивільняється з гранул цих клітин. Симптоми алергії або анафілаксії можна усунути введенням антигістамінних препаратів. У нормальних умовах такі реакції гіперчутливості, які відбуваються за участю тканинних базофілів, мають тенденцію до самообмеження внаслідок виділення цими клітинами хемотоксичного фактора залучення еозинофілів. Ферменти еозинофілів гістаміназа, арилсульфатаза руйнують речовини, які вивільняють тканинні базофіли під час імунних реакцій.
Відомо, що тканинні базофіли походять від стовбурової кровотворної клітини. Недиференційовані попередники тучних клітин мігрують через кров у сполучну тканину, де проліферують і диференціюються у зрілі клітини. У цих процесах беруть участь Т-лімфоцити. Мітотичний поділ тучних клітин спостерігається досить рідко. Оскільки є дані про здатність тканинних базофілів до синтезу ДНК, то можливо мітози трапляються у них частіше, але їх важко побачити через велику кількість гранул, які містяться у цитоплазмі цих клітин.
Адипоцити (жирові клітини). Раніше вони називались ліпоцитами. Ці клітини здатні накопичувати у своїй цитоплазмі резервний жир, який має значення у трофіці, енергоутворенні та метаболізмі води. У пухкій сполучній тканині вони розміщуються групами, рідше поодинці, і, здебільшого, біля кровоносних судин. Коли їх накопичується велика кількість, вони утворюють жирову тканину.
Форма поодинокого адипоцита куляста, а коли їх багато, вони тиснуть один на одного і набувають багатокутної форми. Зріла жирова клітина містить одну велику краплю жиру, яка розтягує всю клітину, так що цитоплазма лише тонким шаром оточує жир. Ядро змінює свою форму, стає сплющеним. Діаметр жирової клітини може досягати 120 мкм. Така клітина на поперечному зрізі подібна до персня з печаткою: ядро — це печатка, а перстень — тонкий шар цитоплазми, що оточує жир. Ліпіди добре забарвлюються Суданом III в оранжевий колір або осмієвою кислотою в чорний колір.
Органели розташовані головним чином навколо ядра, де більше цитоплазми. У жировій клітині є вільні рибосоми, обидва типи ендоплазматичної сітки, комплекс Гольджі та мітохондрії. Скупчення таких жирових клітин утворює білу жирову тканину.
Жирові крапельки, що потрапляють у лімфу, а потім у кров з епітеліоцитів тонкої кишки, розмірами близько 1 мкм мають назву хіломікронів (від грецького «хілос» — сік, «мікрон» — малий).У цих частинках містяться тригліцериди, а також фосфоліпіди, ефір холестерину і деяка кількість білків, які утворюють з ліпідами ліпопротеїни. Під дією ферментів ліпопротеїнліпаз, що виробляє ендотелій судин, тригліцериди хіломікронів розщеплюються на жирні кислоти і гліцерин, які можуть поглинатися жировою клітиною. Під дією гліцерокінази, яка утворюється у цій клітині в процесі вуглеводного обміну, з жирних кислот і гліцерину ресинтезуються тригліцериди. Депонований в адипоцитах жир метаболізує під дією ліполітичних гормонів (адреналін, інсулін), і тканинного ферменту ліпази, який розщеплює тригліцериди до гліцерину і жирних кислот. Останні зв'язуються з альбуміном крові і транспортуються до інших тканин, яким потрібні поживні речовини.
За походженням жирові клітини, очевидно, є окремою клітинною лінією. Жирові клітини живуть довго. Мітози у клітинах-попередниках адипоцитів закінчуються через два-три тижні після народження. У дорослих жирові клітини не діляться, але є дані про те, що нові адипоцити у дорослих можуть утворитися з адвентиційних клітин шляхом накопичення в них жиру.
Пігментоцити (пігментні клітини, меланоцити)містять у своїй цитоплазмі пігмент меланін. Трапляються не лише у сполучній тканині, але й у складі епітеліїв, зокрема, базальному шарі епідермісу. Меланоцити сполучної тканини, як правило, не продукують меланін, а лише фагоцитують його (про що свідчить від'ємна ДОФА-реакція). Єдине виключення — люди монголоїдного типу, у них у сполучній тканині дерми куприкової ділянки трапляються меланінсинтезуючі пігментні клітини, які формують тут так звану монгольську пляму. Меланоцити, на відміну від інших клітинних популяцій сполучної тканини, походять з клітин нервового гребеня, а не з мезенхіми.
Адвентиційні клітини— це популяція малоспеціалізованих клітин, що розташовуються вздовж кровоносних судин. Вони мають плоску або веретеноподібну форму, слабо базофільну цитоплазму, овальне ядро і слабо розвинені органели. У процесі диференціації ці клітини можуть, очевидно, перетворюватися у фібробласти й адипоцити. Багато авторів заперечують існування адвентиційних клітин.
Волокнисті структури. Колагенові волокна.У пухкій сполучній тканині колагенові волокна розташовані у різних напрямках і мають вигляд хвилястих, спірально покручених, круглих або плоских тяжів товщиною 1...10 мкм. Вони здатні утворювати пучки, товщина яких може досягати 150 мкм. У нативному вигляді колагенові волокна безбарвні, на гістологічному препараті забарвлюються оксифільно, при імпрегнації сріблом набирають бурувато-жовтого кольору. Ці волокна не галузяться і не анастомозують між собою.
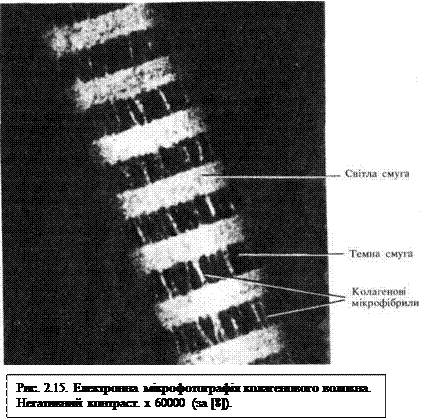
Колагенове волокно (рис. 2.14, рис. 2.15) побудоване з пучків фібрил, зцементованих глікозаміногліканами та глікопротеїнами. Товщина фібрил 50...100 нм. Фібрили мають характерну поперечну посмугованість у вигляді світлих та темних смуг, що чергуються між собою з періодом повторюваності 64 нм. Фібрили складаються з мікрофібрил товщиною близько 10 нм, їх можна побачити в електронному мікроскопі у вигляді ледь хвилястих ниток. Мікрофібрили побудовані із ще тонших елементів – протофібрил, а останні — з молекул колагену. Молекули білка колагену мають довжину близько 280 нм і товщину 1,4 нм. Вони побудовані з трьох поліпептидних ланцюжків попередника колагену — проколагену. Синтез колагену, а також глікозаміногліканів та глікопротеїнів відбувається у клітинах пухкої сполучної тканини — фібробластах. Далі ці речовини секретуються у міжклітинну речовину. Поза клітиною з молекул колагену утворюються протофібрили і т. д. Маркерними амінокислотами зрілого колагену є гідроксипролін та гідроксилізин.
Існує 12 типів колагену, які розрізняють за молекулярною організацією, органною та тканинною належністю. Колаген І типу є в сполучній тканині шкіри, кістки, у рогівці ока, склері, стінці артерій тощо; II типу — у гіаліновому і фіброзному хрящах, у скловидному тілі; III типу — у дермі шкіри плода, в стінці великих кровоносних судин, у ретикулярних волокнах; IV типу — у базальних мембранах, капсулі кришталика; V типу — навколо клітин, що його синтезують у вигляді екзоцитоскелету. Колагени VI, VII типів називають мікрофібрилярними; колагени VIII, IX, X, XI типів — так звані малі, знайдені у невеликих кількостях в ендотелії, хрящах, скловидному тілі.
 Колагенові волокна містять 65 % води. Вони здатні притягати воду і набрякати як у складі організму, так і поза ним. У проточній воді їхня товщина збільшується на 50 % внаслідок набряку, а в підкисленому середовищі — у 500 разів; довжина волокон при цьому не зростає. Такі властивості колагенових волокон зумовлюють їхню функцію в організмі — бути депо води. Цією властивістю колагенових волокон зумовлена поява набряків при патології. При втраті крові вони віддають воду, відновлюючи об'єм крові. При виварюванні колагенові волокна утворюють клей (звідси походить їхня назва «кола» — клей, «гоно» — народжую, продукую). Нони мають невелику резистентністьт до дії кислот, лугів та протеолітичних ферментів. Колагенові волокна дуже міцні, але мають низьку еластичність, їхній модуль пружності 60...70 кг/мм2. Це иайміцніші структури в організмі, основна їхня функція — оморно-механічна.
Колагенові волокна містять 65 % води. Вони здатні притягати воду і набрякати як у складі організму, так і поза ним. У проточній воді їхня товщина збільшується на 50 % внаслідок набряку, а в підкисленому середовищі — у 500 разів; довжина волокон при цьому не зростає. Такі властивості колагенових волокон зумовлюють їхню функцію в організмі — бути депо води. Цією властивістю колагенових волокон зумовлена поява набряків при патології. При втраті крові вони віддають воду, відновлюючи об'єм крові. При виварюванні колагенові волокна утворюють клей (звідси походить їхня назва «кола» — клей, «гоно» — народжую, продукую). Нони мають невелику резистентністьт до дії кислот, лугів та протеолітичних ферментів. Колагенові волокна дуже міцні, але мають низьку еластичність, їхній модуль пружності 60...70 кг/мм2. Це иайміцніші структури в організмі, основна їхня функція — оморно-механічна.
Еластичні волокна, на відміну під колагенових, мають у нативному вигляді жовтуватий колір, галузяться і анастомозують між собою, завжди розташовані поодинці, не утворюють пучків. Товщина їх від 0,3 до 10... 18 мкм.
Основним хімічним складником еластичних волокон є глобулярний білок еластин, який синтезують фібробласти. В еластині міститься велика кількість амінокислот проліну та гліцину, відсутній цистин. Крім того, характерна наявність двох похідних амінокислот — десмозину та ізодесмозину, що зумовлюють його еластичність. Молекули еластину мають форму глобул діаметром 2,8 нм. Поза клітиною вони з'єднуються у ланцюжки товщиною З...3,5 нм, які називаються еластиновими протофібрилами, що в комплексі з глікопротеїнами, утворюють мікрофібрили товщиною 8...10 нм. Пластичне волокно за даними електронної мікроскопії побудоване і двох компонентів — у центрі міститься аморфний компонент, а на периферії — мікрофібрилярний. У різних типах еластичних волокон співвідношення цих двох компонентів різне. Найбільш зрілі еластичні волокна містять близько 90 % еластину у вигляді аморфного компоненту. Мікрофібрилярний компонент сильніше розвинений там, де вимоги до механічної міцності більші, ніж до еластичності. Крім зрілих еластичних волокон, розрізняють близькі до них, так звані елаунінові та окситаланові волокна. В елаунінових волокнах співвідношення мікрофібрил і аморфного компоненту приблизно рівне, а окситаланові складаються лише з мікрофібрил.
Еластичні волокна бідніші водою порівняно з колагеновими (містять 47 % води). Вони стійкі до кип’ятіння, дії кислот, лугів, мацерації, гниття, довше зберігаються у трупному матеріалі, їхня міцність набагато менша, ніж у колагенових волокон, але їм властива висока еластичність. Це прекрасні амортизатори, які забезпечують повернення структур до вихідного положення. З віком еластичність цих волокон знижується, вони розпадаються на фрагменти. Еластичні волокна погано сприймають гістологічні барвники загального характеру, їх можна виявити елективно за допомогою орсеїну або резорцин-фуксину.
 Ретикулярні волокна можна спостерігати у препаратах, імпрегнованих сріблом, тому їх називають ще аргірофільними. Серед останніх розрізняють два типи волокон: власне ретикулярні — це дефінітивні утвори, які побудовані з колагену III типу; преколагенові — початкова стадія при утворенні колагенових волокон у період ембріогенезу, а також при регенерації.
Ретикулярні волокна можна спостерігати у препаратах, імпрегнованих сріблом, тому їх називають ще аргірофільними. Серед останніх розрізняють два типи волокон: власне ретикулярні — це дефінітивні утвори, які побудовані з колагену III типу; преколагенові — початкова стадія при утворенні колагенових волокон у період ембріогенезу, а також при регенерації.
Ретикулярні волокна дуже близькі до колагенових за своїм складом, але відрізняються від них меншою товщиною, розгалуженістю та наявністю анастомозів. Ретикулярні волокна разом з ретикулярними клітинами, що їх продукують, утворюють сітчасті структури.
Електронно-мікроскопічно у ретикулярних волокнах спостерігаються протофібрили товщиною 40 нм, склеєні аморфною речовиною. Протофібрили мають не завжди чітку посмугованість з періодом 64...67 нм (тобто ідентичну колагеновим волокнам). На відміну від колагенових волокон, ретикулярні мають високу концентрацію ліпідів, вуглеводів та сірки. Вони стійкі до дії слабких кислот і лугів, трипсину. За здатністю до розтягання вони займають проміжне положення між колагеновими та еластичними.
Основна міжклітинна речовина (аморфний компонент).Клітини та волокна сполучної тканини занурені в основну речовину, або аморфний компонент. Основна речовина в організмі становить близько 20 % маси тіла. У дитячому віці її більше, ніж у дорослої людини або у старих людей.
Вміст основної речовини неоднаковий у різних видів сполучної тканини. За фізико-хімічним станом це гель непостійної в'язкості та хімічного складу. В утворенні основної речовини беруть участь клітини сполучної тканини і, в першу чергу, фібробласти. Хімічний склад аморфного компоненту характеризується наявністю води, білків, ліпідів, полісахаридів, мінеральних речовин. Вміст полісахаридів 0,5...5 %. До них належать глікозаміноглікани (ГАГ): сульфатовані — гепарансульфат, хондроїтин-4-сульфат, хондроїтин-6-сульфат, дерматансульфат, а також несульфатовані, представником яких є гіалуронова кислота. Сульфатовані ГАГ сполучаються з білками, утворюючи протеоглікани. Глікозаміноглікани зумовлюють консистенцію аморфної речовини та її функціональні властивості. У свою чергу, функціональні риси сполучної тканини загалом залежать від фізико-хімічного стану основної речовини. Чим вона щільніша, тим більше виражена механічна, опорна функція. Рідша за консистенцією основна речовина краще забезпечує трофічну функцію. Гістамін і гіалуронідаза збільшують проникливість аморфного компоненту (є багато мікроорганізмів, споряджених гіалуронідазою, яка допомагає їм прокладати шлях у сполучній тканині). Підвищення концентрації ГАГ (зокрема гіалуронової кислоти), навпаки, знижує проникність основної міжклітинної речовини.
Основна речовина створює передумови для пересування клітин, здатних до руху, є шляхом транспорту поживних речовин і продуктів метаболізму.
Щільна волокниста сполучна тканина (textus connectives fibrosus compactus).Для цього виду сполучної тканини характерним є переважний розвиток волокнистих структур і, у першу чергу, колагенових волокон. Ця особливість забезпечує високі амортизаційно-механічні властивості. Залежно від способу орієнтації колагенових волокон у просторі розрізняють оформлену щільну волокнисту сполучну тканину і неоформлену щільну волокнисту сполучну тканину.
Оформлена щільна волокниста сполучна тканина розміщена у складі фіброзних мембран, зв'язок, сухожиль. Останні, з'єднуючи м'язи з кістками, зазнають дії вектора сили переважно в одному напрямі. Вказаний фактор є причиною строго паралельної орієнтації пучків колагенових волокон у просторі. Між окремими пучками волокон розміщені високодиференційовані клітини фібробластичного ряду (фіброцити), які у результаті своєї синтетичної діяльності забезпечують фізіологічну регенерацію сухожильних пучків. Пучок колагенових волокон, оточений шаром фіброцитів, називається сухожильним пучком першого порядку (рис. 2.16). Фіброцити розмежовують сусідні сухожильні пучки першого порядку, і на поздовжньому розрізі сухожилля мають вигляд рисочок. Характерним є чергування пучків колагенових волокон і рядів фіброцитів. На поперечному розрізі сухожилля можна побачити характерні пластинчасті відростки фіброцитів, які вини  кають внаслідок стискання клітинного тіла сусідніми колагеновими волокнами. Кілька сухожильних пучків першого порядку в своїй сукупності утворюють сухожильні пучки другого порядку, останні розмежовані прошарками пухкої сполучної тканини, що носять назву ендотендинію. У складі великих сухожиль пучки другого порядку, об'єднуючись, утворюють сухожильні пучки третього і навіть четвертого порядків. Зовні сухожилля оточене перитендинієм, утвореним пухкою сполучною тканиною.
кають внаслідок стискання клітинного тіла сусідніми колагеновими волокнами. Кілька сухожильних пучків першого порядку в своїй сукупності утворюють сухожильні пучки другого порядку, останні розмежовані прошарками пухкої сполучної тканини, що носять назву ендотендинію. У складі великих сухожиль пучки другого порядку, об'єднуючись, утворюють сухожильні пучки третього і навіть четвертого порядків. Зовні сухожилля оточене перитендинієм, утвореним пухкою сполучною тканиною.
Прикладом неоформленої щільної волокнистої сполучної тканини може служити сітчастий шар дерми шкіри. У його складі товсті пучки колагенових волокон ідуть у різних напрямах, що забезпечує резистентність шкіри при найрізноманітніших напрямках дії механічних факторів. Між пучками колагенових волокон розміщені фібробласти та макрофаги, судинно-нервові пучки та основна міжклітинна речовина.
 Сполучні тканини зі спеціальними властивостями.Для сполучних тканин цієї групи характерний переважний розвиток того чи іншого різновиду клітинних елементів, а також ряд особливостей міжклітинної речовини.
Сполучні тканини зі спеціальними властивостями.Для сполучних тканин цієї групи характерний переважний розвиток того чи іншого різновиду клітинних елементів, а також ряд особливостей міжклітинної речовини.
Жирова тканина (textus adiposus).Характерною її особливістю є переважний розвиток жирових клітин — адипоцитів. Розрізняють два види жирової тканини — білу і буру. Біла жирова тканина побудована з описаних вище адипоцитів, які містять одну велику краплю жиру. Жирові клітини утворюють часточки різних розмірів і форми. Між ними розміщені вузенькі прошарки пухкої сполучної тканини, у якій є фібробласти, тканинні базофіли, лімфоцити, тонкі колагенові волокна. Тут також локалізовані кровоносні та лімфатичні капіляри, які охоплюють своїми петлями жирові часточки. Біла жирова тканина відіграє, в основному, роль депо високоенергетичного поживного матеріалу, яким для організму є нейтральні жири. Вона також бере участь в обміні води, виконує амортизаційні функції при захисті життєво важливих органів від механічних пошкоджень. Білий жир у людини розміщений переважно на передній черевній стінці, на стегнах, у ділянках сідниць, в очеревині, підшкірній жировій клітковині. Під час голодування підшкірна, колониркова жирова тканина, а також сальник швидко втрачають запаси жиру. На відміну від цього, жирова тканина в ділянці долонь і підошв, очної орбіти навіть при тривалому голодуванні майже не втрачає ліпідів, тому що у таких ділянках її основною функцією є механічна, а не обмінна.
Бура жирова тканина складається з адипоцитів, які містять у цитоплазмі велику кількість дрібних жирових включень. Ядро займає центральне положення. У цитоплазмі цих клітин є значна кількість мітохондрій, цитохроми останніх зумовлюють бурий колір тканини. Адипоцити мають високу окисну здатність, а їхній обмін забезпечує вивільнення тепла, яке зігріває кров у численних капілярах між клітинами. Таким чином, основна функція цієї тканини терморегуляторна. Вважають, що бурий жир у людини є лише у дитячому віці; найчастіше він локалізований у міжлопаточній ділянці, на шиї, під пахвами, в колонирковій клітковині. Запаси його у немовлят становлять 30 г. Однак існують дані, що у перинефральних жирових депо, які є основним місцем локалізації цієї тканини у людини, знайдено бурий жир в осіб віком до 50 років.
Ретикулярна тканина (textus reticularis)утворює сполучнотканинну строму кровотворних органів, формуючи мікрооточення для клітин крові, що дозрівають. В основі ретикулярної тканини лежать ретикулярні клітини і волокна. Ретикулярні клітини мають відростки, якими вони контактують одна з одною, утворюючи сітку. Сітка доповнюється ретикулярними волокнами, які тісно пов'язані з клітинами. Серед ретикулярних клітин розрізняють фібробластоподібні клітини, фагоцити моноцитарного генезу та малодиференційовані клітини.
Пігментна тканина (textus pigmentosus)порівняно з іншими видами сполучної тканини збагачена пігментними клітинами — меланоцитами. Пігментної тканини багато у райдужній оболонці ока, у шкірі сосків молочних залоз, навколо анального отвору. Пігментні клітини у зв'язку з високим вмістом меланіну, який може поглинати ультрафіолетові промені, відіграють захисну роль щодо пошкоджуючої дії сонячної радіації.
Слизова тканина (textus mucosus)розміщена у складі пупкового канатика зародка. її особливість — відсутність волокнистих структур і значний вміст в основній міжклітинній речовині високомолекулярних біополімерів, які забезпечують тургор (пружність) тканин пупкового канатика і попереджують можливість перетискання кровоносних судин, що живлять зародок.
Дата добавления: 2014-12-03; просмотров: 780; Мы поможем в написании вашей работы!; Нарушение авторских прав |