КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Систематичне положення людини
Питання про таксономічне положення людини вперше було поставлене на наукову основу в середині 18ст., коли К. Ліней включив людину в систему тваринного світу.Він об'єднав людей з мавпами в один ряд приматів.Пізніше питаннями систематики людини займалисяР. Гекслі, Е.Фогт, Ч. Дарвін і багато інших науковців.
У наш час розроблено декілька класифікацій ряду приматів. Ми пропонуємо одну з них (виділяється лінія, що веде до людини): домен -евкаріоти, надцарство -задньоджутикові, царство – тварини, підцарство - багатоклітинні, грантип- білатеральносиметричні, надтип - вториннороті, тип – хордові,підтип – хребетні, інфратип -щелепнороті,надклас -четвероногі, клада -амніоти, інфраклада -синапсиди, клас– ссавці, підклас - живородні, інфраклас - голотерії, н/легіон -технотерії, легіон -кладотерії, підлегіон -затерії, інф/легіон -трібосфеніди, надкогорта -терії, або справжні звірі, когорта -плацентарні, підкогорта - ексафриканські звірі, магнряд- північні звірі, надряд -еуархонтогліреси, гранряд –першопредки (еуархонти), мірряд - приматоподібні, ряд– примати, що вміщує два підряди- мокроносі та сухоносі примати. У підряді сухоносих мавп виділяють 2 інфраряди:довгопятові (тарзіоморфи)та антропоїди (мавпи).У інфраряді антропоїдів виділяють двапарвряди:широконосі (близько 50 видів, що у тропічних регіонах Нового Світу) та вузьконосі (100 видів мавп, які населяють тропіки Старого Світу).Серед вузьконосих мавп виділяють3 надродини:тонкотілі мавпи, собакоголові та гоміноїди. В надродині гоміноїдів є2 родини:малі людиноподібні (гіббони) та великі людиноподібні мавпи. Великі людиноподібні мавпи поділяються на 2 підродини : великі азійські мавпи (2 види орангутан) та великі африканські мавпи - гомініни(горила, шимпанзе, людина). Підродина гомінінвключає кілька родів - шимпанзе, сахелантроп, орорін, ардіпітек, австралопітек, Homo або людина. До складу роду людина входить 1 вид- Homo sapiens або людина розумна. Вид має 2 підвиди: людина розумна давня та людина розумна розумна.


Еволюція ранніх ссавців.
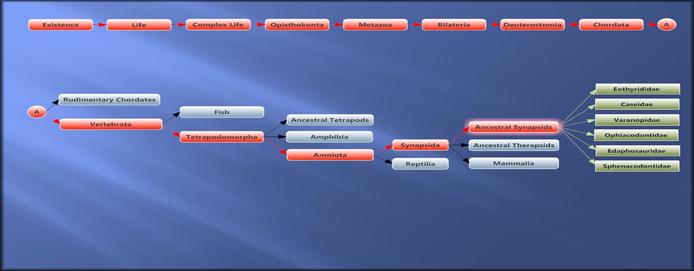
Схема еволюції синапсід. Бічні гілки примитивних синапсід.

Схема еволюції терапсід.
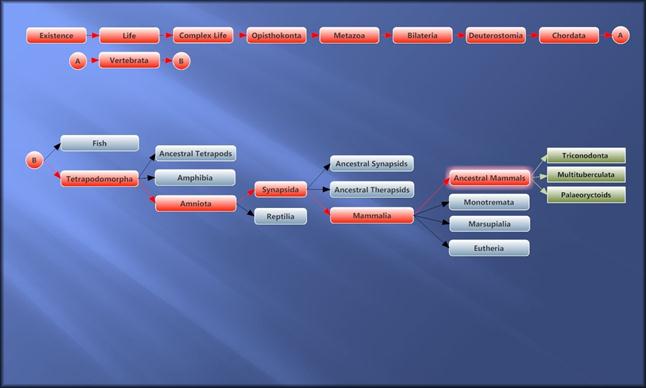
Схема еволюції маммалій.

Перший етап еволюції синапсид (формотворення).

Другий етап еволюції синапсид (формотворення терапсід).

Третій етап еволюції синапсид (формотворення теріодонтів і маммалій).

Аделобазилевс - форма, що відділилася від загальних предків звірів та інших цинодонтів (225 млн. р. т.).

Мегазостродон - примитивний теріоморф (200 млн. р. т.), морфологічно близький до звірів.
1.3. I етап еволюції приматів
Усі особливості еволюційного розвитку приматів пов'язані зі зміною навколишнього середовища, тобто із геолого-кліматичними умовами, котрі складалися у певні геологічні періоди на Землі. Відповідно до них виділяють три етапи філогенетичного розвиткуприматів.
Перший етап еволюції приматів
Геолого - кліматичні зміни:два доісторичних континенти (північний - Лавразія і південний – Гондвана) розкололися на шматки. Материки почали набувати сучасних обрисів:Північна Америка і Європа відокремилися одна від одної, аПівденна Америка перетворилася на острів поруч з Африкою.Клімат ставав усе більш континентальним, вимирали динозаври, звільнилося багато екологічних ніш. Це викликалолавиноподібне формоутворення серед тих груп тварин, які виявилися краще пристованими до нових умов існування.
На початку мезозойської ери (тріас, 225 млн років тому) від цинодонтів, однієї з гілок розвитку синапсид, які ще зберігали багато спільних рис з амфібіями, виділилися групи докодонтів, потім трикодонтів, гобіконодонтів, австралофенід (однопроходні ссавці Австралії) і трибугорчастих ссавців (поетапно - симметродонти,дріолестіди, бореосфеніди, а також пантотерії, або справжні звірі), які в наступному Юрському періоді дали початок першим плацентарним - еомайя, юрамайя, акрістатеріум (160-172 млн. р.т.) і сумчастим ссавцям відповідно. Вони відрізнялися недостатньою дифференціацією зубів (усі бугорчасті).
 Розвиток щелеп та вушних кісточок терапсид.
Розвиток щелеп та вушних кісточок терапсид.
 Розвиток щелеп та зубів маммалій.
Головний мозок ссавців, незважаючи на збереження загального типу організаціїї з циркулярними борознами і звивинами, має багато своєрідних рис, котрі зумовлені специфікою еволюції окремих систематичних груп. У багатьох видів тварин кора має тільки ядерні зони аналізаторів, тобто клітинні поля первинного кіркового аналізу. Поява форм із більш розвиненим мозком ( у корі формуються поля вторинного кіркового аналізу) призвела до його суттєвої перебудови: дуже зменшується площа палеокортексу і нюхового мозку, збільшуються розміри неокортексу ( спочатку зникають борозни і зростають розміри мозку, а потім утворюється система нових радіальних борозен на межіфункціональних областей). Це був новий напрям еволюції головного мозку, який почав складатися у комахоїдних ссавців, аз часом став характерним і для приматів. Усі еволюційні етапи подібних перетворень віднайдені на ендокранах викопних тварин від лемурів до великих людиноподібних мавп.
Перші ссавці.
Розвиток щелеп та зубів маммалій.
Головний мозок ссавців, незважаючи на збереження загального типу організаціїї з циркулярними борознами і звивинами, має багато своєрідних рис, котрі зумовлені специфікою еволюції окремих систематичних груп. У багатьох видів тварин кора має тільки ядерні зони аналізаторів, тобто клітинні поля первинного кіркового аналізу. Поява форм із більш розвиненим мозком ( у корі формуються поля вторинного кіркового аналізу) призвела до його суттєвої перебудови: дуже зменшується площа палеокортексу і нюхового мозку, збільшуються розміри неокортексу ( спочатку зникають борозни і зростають розміри мозку, а потім утворюється система нових радіальних борозен на межіфункціональних областей). Це був новий напрям еволюції головного мозку, який почав складатися у комахоїдних ссавців, аз часом став характерним і для приматів. Усі еволюційні етапи подібних перетворень віднайдені на ендокранах викопних тварин від лемурів до великих людиноподібних мавп.
Перші ссавці.
 Моргауконодон.
Моргауконодон.
 Представник триконодонтів - гобіконодон.
Представник триконодонтів - гобіконодон.
 Триконодон - янодон.
Ознаки ссавців:
· живородіння (крім представників підкласу першозвірів);
· справжня гомойотермія (теплокровність; виключенням є гризун голий землекоп);
· наявність волосяного покрива (шерсті), сильний развиток шкірних залоз (потових та сальних), а також рогових утворень;
· наявність в черепі однієї скулової дуги;
· наявність чіткого розчленування хребта на 5 відділів (шийний, грудний, поясничний, скрижевий, хвостовий);
· платіцільний тип хребців;
· наявність 7 хребців в шийному відділі хребта (як виключення у трипалих лінивців їх 9, а у лінивця Гофмана — 6), до першого із яких двома потиличними мищелками приєднується череп;
· наявність діафрагми, развиток підшкірної мускулатури;
· високий рівень розвитку нервової системи, який забезпечує гнучке реагування на вплив з боку зовнішнього середовища;
· особливий тип будови головного мозка (в тому числі сильний розвиток переднього мозку, перехід до нього функцій основного зорового центру та центру керування складними формами поведінки);
· специфічний характер залягання сірої речовини в спинному мозкові, при якому воно утворює при поперечному перетині фігуру «метелика» з бічними виступами — «рогами»;
· наявність пірамідального шляху від переднього мозка до продовгуватого таспинного
; · взаємне накладання областей іннерваціїсимпатичної та парасимпатичної нервових систем впритул до повного перекривання;
· наявність трьох слухових кісточок середнього вуха, зовнішнього вушного проходу та вушної раковини;
· альвеолярна будова легень;
· чотирикамерне серце;
· наявність однієї (лівої) дуги аорти;
· відсутність воротного кровообігу в нирках, характерного для решти четвероногих;
· безядерні еритроцити;
· гетеродонтні (диференційовані на різці, ікла, передкорінні та корінні) зуби, сидячі в ячейках (альвеолах) щелеп;
· наявність у більшості ссавців значної кількості лімфатичної тканини в стінках слепої кишки;
· овальна форма сімянників;
· схильність до пріонних захворювань (крім ссавців, вони відомі лише у деяких грибів).
Перші плацентарні
Триконодон - янодон.
Ознаки ссавців:
· живородіння (крім представників підкласу першозвірів);
· справжня гомойотермія (теплокровність; виключенням є гризун голий землекоп);
· наявність волосяного покрива (шерсті), сильний развиток шкірних залоз (потових та сальних), а також рогових утворень;
· наявність в черепі однієї скулової дуги;
· наявність чіткого розчленування хребта на 5 відділів (шийний, грудний, поясничний, скрижевий, хвостовий);
· платіцільний тип хребців;
· наявність 7 хребців в шийному відділі хребта (як виключення у трипалих лінивців їх 9, а у лінивця Гофмана — 6), до першого із яких двома потиличними мищелками приєднується череп;
· наявність діафрагми, развиток підшкірної мускулатури;
· високий рівень розвитку нервової системи, який забезпечує гнучке реагування на вплив з боку зовнішнього середовища;
· особливий тип будови головного мозка (в тому числі сильний розвиток переднього мозку, перехід до нього функцій основного зорового центру та центру керування складними формами поведінки);
· специфічний характер залягання сірої речовини в спинному мозкові, при якому воно утворює при поперечному перетині фігуру «метелика» з бічними виступами — «рогами»;
· наявність пірамідального шляху від переднього мозка до продовгуватого таспинного
; · взаємне накладання областей іннерваціїсимпатичної та парасимпатичної нервових систем впритул до повного перекривання;
· наявність трьох слухових кісточок середнього вуха, зовнішнього вушного проходу та вушної раковини;
· альвеолярна будова легень;
· чотирикамерне серце;
· наявність однієї (лівої) дуги аорти;
· відсутність воротного кровообігу в нирках, характерного для решти четвероногих;
· безядерні еритроцити;
· гетеродонтні (диференційовані на різці, ікла, передкорінні та корінні) зуби, сидячі в ячейках (альвеолах) щелеп;
· наявність у більшості ссавців значної кількості лімфатичної тканини в стінках слепої кишки;
· овальна форма сімянників;
· схильність до пріонних захворювань (крім ссавців, вони відомі лише у деяких грибів).
Перші плацентарні
 Еомайя - один з найперших плацентарних ссавців (135 млн. р.т.).
Еомайя - один з найперших плацентарних ссавців (135 млн. р.т.).
 Юрамайя - перше плацентарне (160 млн.р.т.).
Відокремлення перших приматів з групи приматоморфних ссавців (предки кагуан та приматів) відбулося приблизно 79 млн років тому, у другій половині крейдяного періоду. Перед цим від загальних предків відєдналися предки тупай (86, 2 млн. р. т.), гризуни з зайцеподібними (85-100 млн. р. т.) та інші бореотерії - лавразитерії (понад 100 млн. р. т.). Одним із перших був пургаторіус. Ці тварини були розміром від миші до кішки, а способом пересування нагадували білку. Близько 65 млн років тому значно зрослозагальне різноманіття ссавців, виникли копитні травоїдні, першіхижаки та інші види тварин.
На початку еоцену (приблизно 55 млн років тому) суттєво збільшилася конкуренція і гризуни витиснули перших приматів із землі на дерева.Та частина ранніх приматів, яка зоставалася у звичних умовах існування протягом тривалого часу і не зазнала істотних перетворень, започаткувала підряд напівмавп. Характерні ознаки справжніх приматів формувалися упродовж кайнозойської ери як пристосування до життя на деревах.
Особливості ряду приматів
Унаслідок деревного способу життя п'ятипала кінцівка хребетних у мавп стає більш рухливою. Добре розвивається здатність до захоплювання і утримання предметів. Кігті замінються на нігті, а подушечки пальців стають органами дотику (в інших ссавців органами дотику слугують вібриси). Верхні кінцівки набувають здатності до обертальних рухів у ліктьовому суглобі, а добре розвинений плечовий пояс призводить до формування такого специфічного для приматів способу локомоції, як брахіація, тобто пересування за допомогою рук при висячому положенні тіла. Більшість мавп можуть сидіти, стрибати або висіти, підтримуючи тіло в більш-менш випрямленому положенні. Це зміцнювало пояс нижніх кінцівок і дозволило в майбутньому деяким групам приматів перейти до прямоходіння.
Мавпи, як і люди,мають досить несприятливе співвідношення маси і поверхні тіла в тому розумінні, що надмірна вага може спричинити серйозні наслідки навіть при незначних падіннях. При житті на деревах ці особливості могли стати джерелом підвищеного рівня травматизму та загибелі особин. За таких умов виживання приматів було можливим лише внаслідок формування ефективної системи зворотних зв'язків між кінцівками і відповідними відділами центральної нервової системи. Це призводило до прогресивної еволюції головного мозку і нової кори, тобто до структурної церебралізації. Подальший розвиток одержали зоровий і слуховий аналізатори, відбувалася певна редукція нюху. Для мавп характерними є великі очі, високодиференційована сітківка, кольоровий і стереоскопічний зір. Остання властивість пов'язана з тим, що очі приматів розташовані в одній площині, а не з боків голови, як у багатьох ссавців і нижчих мавп, наприклад у лемурів. Крім того, приблизно половина волокон зорового тракту перехрещуються на рівні гіпоталамусу проміжного мозку, що призводить до накладення полів зору і стереоскопічного ефекту. Унаслідок подібних морфофізіологічних перебудов у приматів розвиваються такі нові якості інтеграції, як об'ємно-просторово-оптичне сприйняття; синтез у сприйнятті просторових співвідношень об'єктів; формування структурних гомологів тих ділянок головного мозку, з якими пов'язані вищі психічні функції людини (концептуальне мислення, членороздільна мова, трудова діяльність тощо).
Особливостями життя на деревах пояснюється і специфічна дієта приматів. Вони, головним чином, рослинноїдні, рідше – комахоїдні або всеїдні.
Розмноження перестає бути сезонним і примати переходять до поліциклічності, а термін вагітності збільшується до 9 місяців (шимпанзе, горила). Завдяки розвитку вищої нервової діяльності і соціальних відносин в угруповуваннях приматів збільшується тривалість життя окремих особин, а період дитинства зростає до 8-12 років.
Подібні перетворення становлять собою складову частину загальнобіологічних еволюційних змін онтогенезу, які характерні для усіх організмів. У ході еволюції здійснюється перехід від запліднення і розвитку організмів у зовнішньому середовищі до розвитку під захистом яйцевих оболонок, а потім і до внутрішньоутробного розвитку. Одночасно різко зменшується кількість гамет і плодючість, але суттєво зростає виживання окремих особин завдяки розвиненій турботі про потомство і зростанню ролі навчання. Подібні процеси взагалі характерні для прогресивної еволюції індивідуального розвитку вищих тварин. Вони спрямовані на збільшення його незалежності від середовища існування і називаються автономізацією онтогенезу.
Для більшості приматів характерним є стадний, груповий спосіб життя. Він пов'язаний з формуванням складної системи комунікації (міміка, вокалізація). Сучасні мавпи добре засвоюють мову жестів – амслен. Розвиваються орієнтовно-дослідницька, маніпулятивна і наслідувальна діяльності. Це призводить до того, що індивідуальні досягнення кожної особини швидко стають колективними і суттєво збільшують шанси на виживання усієї групи.
Таким чином, у приматів, внаслідок життя на деревах, сформувалися певні особливості анатомічної будови, психічного відображення і соціальних взаємовідносин. Наступний геологічний період в історії Землі (еоцен) пов'язаний з розквітом цієї адаптивно пластичної групи ссавців.
Юрамайя - перше плацентарне (160 млн.р.т.).
Відокремлення перших приматів з групи приматоморфних ссавців (предки кагуан та приматів) відбулося приблизно 79 млн років тому, у другій половині крейдяного періоду. Перед цим від загальних предків відєдналися предки тупай (86, 2 млн. р. т.), гризуни з зайцеподібними (85-100 млн. р. т.) та інші бореотерії - лавразитерії (понад 100 млн. р. т.). Одним із перших був пургаторіус. Ці тварини були розміром від миші до кішки, а способом пересування нагадували білку. Близько 65 млн років тому значно зрослозагальне різноманіття ссавців, виникли копитні травоїдні, першіхижаки та інші види тварин.
На початку еоцену (приблизно 55 млн років тому) суттєво збільшилася конкуренція і гризуни витиснули перших приматів із землі на дерева.Та частина ранніх приматів, яка зоставалася у звичних умовах існування протягом тривалого часу і не зазнала істотних перетворень, започаткувала підряд напівмавп. Характерні ознаки справжніх приматів формувалися упродовж кайнозойської ери як пристосування до життя на деревах.
Особливості ряду приматів
Унаслідок деревного способу життя п'ятипала кінцівка хребетних у мавп стає більш рухливою. Добре розвивається здатність до захоплювання і утримання предметів. Кігті замінються на нігті, а подушечки пальців стають органами дотику (в інших ссавців органами дотику слугують вібриси). Верхні кінцівки набувають здатності до обертальних рухів у ліктьовому суглобі, а добре розвинений плечовий пояс призводить до формування такого специфічного для приматів способу локомоції, як брахіація, тобто пересування за допомогою рук при висячому положенні тіла. Більшість мавп можуть сидіти, стрибати або висіти, підтримуючи тіло в більш-менш випрямленому положенні. Це зміцнювало пояс нижніх кінцівок і дозволило в майбутньому деяким групам приматів перейти до прямоходіння.
Мавпи, як і люди,мають досить несприятливе співвідношення маси і поверхні тіла в тому розумінні, що надмірна вага може спричинити серйозні наслідки навіть при незначних падіннях. При житті на деревах ці особливості могли стати джерелом підвищеного рівня травматизму та загибелі особин. За таких умов виживання приматів було можливим лише внаслідок формування ефективної системи зворотних зв'язків між кінцівками і відповідними відділами центральної нервової системи. Це призводило до прогресивної еволюції головного мозку і нової кори, тобто до структурної церебралізації. Подальший розвиток одержали зоровий і слуховий аналізатори, відбувалася певна редукція нюху. Для мавп характерними є великі очі, високодиференційована сітківка, кольоровий і стереоскопічний зір. Остання властивість пов'язана з тим, що очі приматів розташовані в одній площині, а не з боків голови, як у багатьох ссавців і нижчих мавп, наприклад у лемурів. Крім того, приблизно половина волокон зорового тракту перехрещуються на рівні гіпоталамусу проміжного мозку, що призводить до накладення полів зору і стереоскопічного ефекту. Унаслідок подібних морфофізіологічних перебудов у приматів розвиваються такі нові якості інтеграції, як об'ємно-просторово-оптичне сприйняття; синтез у сприйнятті просторових співвідношень об'єктів; формування структурних гомологів тих ділянок головного мозку, з якими пов'язані вищі психічні функції людини (концептуальне мислення, членороздільна мова, трудова діяльність тощо).
Особливостями життя на деревах пояснюється і специфічна дієта приматів. Вони, головним чином, рослинноїдні, рідше – комахоїдні або всеїдні.
Розмноження перестає бути сезонним і примати переходять до поліциклічності, а термін вагітності збільшується до 9 місяців (шимпанзе, горила). Завдяки розвитку вищої нервової діяльності і соціальних відносин в угруповуваннях приматів збільшується тривалість життя окремих особин, а період дитинства зростає до 8-12 років.
Подібні перетворення становлять собою складову частину загальнобіологічних еволюційних змін онтогенезу, які характерні для усіх організмів. У ході еволюції здійснюється перехід від запліднення і розвитку організмів у зовнішньому середовищі до розвитку під захистом яйцевих оболонок, а потім і до внутрішньоутробного розвитку. Одночасно різко зменшується кількість гамет і плодючість, але суттєво зростає виживання окремих особин завдяки розвиненій турботі про потомство і зростанню ролі навчання. Подібні процеси взагалі характерні для прогресивної еволюції індивідуального розвитку вищих тварин. Вони спрямовані на збільшення його незалежності від середовища існування і називаються автономізацією онтогенезу.
Для більшості приматів характерним є стадний, груповий спосіб життя. Він пов'язаний з формуванням складної системи комунікації (міміка, вокалізація). Сучасні мавпи добре засвоюють мову жестів – амслен. Розвиваються орієнтовно-дослідницька, маніпулятивна і наслідувальна діяльності. Це призводить до того, що індивідуальні досягнення кожної особини швидко стають колективними і суттєво збільшують шанси на виживання усієї групи.
Таким чином, у приматів, внаслідок життя на деревах, сформувалися певні особливості анатомічної будови, психічного відображення і соціальних взаємовідносин. Наступний геологічний період в історії Землі (еоцен) пов'язаний з розквітом цієї адаптивно пластичної групи ссавців.
 Пургаторіус Пургаторіус  Загальна схема еволюції приматів.
Плезіадапіди
Загальна схема еволюції приматів.
Плезіадапіди
 Плезіадапіс
Плезіадапіс
 Карполестес Симона
Карполестес Симона
 Дріомоміс
Дріомоміс
|
1.4. II етап эволюції приматів
Еоценові примати були представлені такими напівмавпами як адапіди, лемуроїди, омоміміди та тарзіоїди (довгоп'яти).
Адапіди виникли в середині палеоцену та нараховували приблизно 20 родів і до нашого часу всі вимерли. У порівнянні з ранніми приматами в них була коротша мордочка і збільшений головної мозок, очі - великі й спрямовані вперед, на кінцівках - нігті. Жили ці тварини в Північній Америці і Європі. Лемуроїди, які можливо походять від адапід, на відміну від них, мали довгі нижні виступаючі зуби, що перетворилися у своєрідний гребінь для догляду за хутром. Налічувалося близько 30 родів цих приматів, які майже всі вимерли. Відносна розмаїтість лемурів у наш час збереглася тільки на острові Мадагаскар, оскільки на ньому майже відсутні хижаки. Довгоп'яти (тарзіоїди) та їх безпосередні попредники Омоміміди були поширені починаючи з середини палеоцену, але вже в олігоцені більшість із них вимерла і до сьогодні з більше ніж 30 родів збереглося усього 3 види, котрі зустрічаються на островах Борнео та Суматра і ведуть, головним чином, нічний спосіб життя.
Наприкінці еоцену відбувся обмін фаунами між Азією і Північною Америкою, остаточно перервався сухопутний зв'язок між Європою і Північною Америкою. Для еволюції приматів цього часу характерним є існування амфіпітека погаунгського. Ця істота поєднувала у своїй будові ознаки вищих і нижчих приматів.
Адапіди.
 Дарвініус
Дарвініус
 Адапіс
Адапіс
Викопні лемуриди.

Дата добавления: 2014-12-30; просмотров: 1385; Мы поможем в написании вашей работы!; Нарушение авторских прав |