КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Классификация. На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны
На основании числа и расположения дендритов и аксона нейроны делятся на безаксонные, униполярные нейроны, псевдоуниполярные нейроны, биполярные нейроны и мультиполярные (много дендритных стволов, обычно эфферентные) нейроны.
Безаксонные нейроны — небольшие клетки, сгруппированы вблизи спинного мозга в межпозвоночных ганглиях, не имеющие анатомических признаков разделения отростков на дендриты и аксоны. Все отростки у клетки очень похожи. Функциональное назначение безаксонных нейронов слабо изучено.
Униполярные нейроны — нейроны с одним отростком, присутствуют, например в сенсорном ядре тройничного нерва в среднем мозге. Многие морфологи считают, что униполярные нейроны в теле человека и высших позвоночных не встречаются.
Биполярные нейроны — нейроны, имеющие один аксон и один дендрит, расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях.
Мультиполярные нейроны — нейроны с одним аксоном и несколькими дендритами. Данный вид нервных клеток преобладает в центральной нервной системе.
Псевдоуниполярные нейроны — являются уникальными в своём роде. От тела отходит один отросток, который сразу же Т-образно делится. Весь этот единый тракт покрыт миелиновой оболочкой и структурно представляет собой аксон, хотя по одной из ветвей возбуждение идёт не от, а к телу нейрона. Структурно дендритами являются разветвления на конце этого (периферического) отростка. Триггерной зоной является начало этого разветвления (то есть находится вне тела клетки). Такие нейроны встречаются в спинальных ганглиях.
Функциональная классификация[править | править вики-текст]
По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (часть из них называется двигательными нейронами, иногда это не очень точное название распространяется на всю группу эфферентов) и интернейроны (вставочные нейроны).
Афферентные нейроны (чувствительный, сенсорный, рецепторный или центростремительный). К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Эфферентные нейроны (эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
Ассоциативные нейроны (вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Секреторные нейроны — нейроны, секретирующие высокоактивные вещества (нейрогормоны). У них хорошо развит комплекс Гольджи, аксон заканчивается аксовазальными синапсами.
Морфологическая классификация[править | править вики-текст]
Морфологическое строение нейронов многообразно. В связи с этим при классификации нейронов применяют несколько принципов:
· учитывают размеры и форму тела нейрона;
· количество и характер ветвления отростков;
· длину аксона и наличие специализированных оболочек.
По форме клетки, нейроны могут быть сферическими, зернистыми, звездчатыми, пирамидными, грушевидными, веретеновидными, неправильными и т. д. Размер тела нейрона варьирует от 5 мкм у малых зернистых клеток до 120—150 мкм у гигантских пирамидных нейронов.
По количеству отростков выделяют следующие морфологические типы нейронов[1]:
· униполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге;
· псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
· биполярные нейроны (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
· мультиполярные нейроны (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
Развитие и рост нейрона[править | править вики-текст]
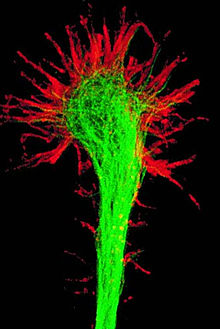
Конус роста
Нейрон развивается из небольшой клетки-предшественницы, которая перестаёт делиться ещё до того, как выпустит свои отростки. (Однако, вопрос о делении нейронов в настоящее время остаётся дискуссионным.) Как правило, первым начинает расти аксон, а дендриты образуются позже. На конце развивающегося отростка нервной клетки появляется утолщение неправильной формы, которое, видимо, и прокладывает путь через окружающую ткань. Это утолщение называется конусом роста нервной клетки. Он состоит из уплощенной части отростка нервной клетки с множеством тонких шипиков. Микрошипики имеют толщину от 0,1 до 0,2 мкм и могут достигать 50 мкм в длину, широкая и плоская область конуса роста имеет ширину и длину около 5 мкм, хотя форма её может изменяться. Промежутки между микрошипиками конуса роста покрыты складчатой мембраной. Микрошипики находятся в постоянном движении — некоторые втягиваются в конус роста, другие удлиняются, отклоняются в разные стороны, прикасаются к субстрату и могут прилипать к нему.
Конус роста заполнен мелкими, иногда соединёнными друг с другом, мембранными пузырьками неправильной формы. Непосредственно под складчатыми участками мембраны и в шипиках находится плотная масса перепутанных актиновых филаментов. Конус роста содержит также митохондрии, микротрубочки и нейрофиламенты, аналогичные имеющимся в теле нейрона.
Вероятно, микротрубочки и нейрофиламенты удлиняются главным образом за счёт добавления вновь синтезированных субъединиц у основания отростка нейрона. Они продвигаются со скоростью около миллиметра в сутки, что соответствует скорости медленного аксонного транспорта в зрелом нейроне. Поскольку примерно такова и средняя скорость продвижения конуса роста, возможно, что во время роста отростка нейрона в его дальнем конце не происходит ни сборки, ни разрушения микротрубочек и нейрофиламентов. Новый мембранный материал добавляется, видимо, у окончания. Конус роста — это область быстрогоэкзоцитоза и эндоцитоза, о чём свидетельствует множество находящихся здесь пузырьков. Мелкие мембранные пузырьки переносятся по отростку нейрона от тела клетки к конусу роста с потоком быстрого аксонного транспорта. Мембранный материал, видимо, синтезируется в теле нейрона, переносится к конусу роста в виде пузырьков и включается здесь в плазматическую мембрану путём экзоцитоза, удлиняя таким образом отросток нервной клетки.
Росту аксонов и дендритов обычно предшествует фаза миграции нейронов, когда незрелые нейроны расселяются и находят себе постоянное место.
Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в среднем в 10-50 раз больше, чем нейронов. Термин ввёл в 1846 году Рудольф Вирхов[1].
Глиальные клетки имеют общие функции и, частично, происхождение (исключение — микроглия). Они составляют специфическое микроокружение для нейронов, обеспечивая условия для генерации и передачи нервных импульсов, а также осуществляя часть метаболических процессов самого нейрона.
Нейроглия выполняет опорную, трофическую, секреторную, разграничительную и защитную функции.
11. Слуховой анализатор. Звукоулавливающий и звукопроводящий отделы слухового анализатора. Строение улитки, микроструктура кортиева органа. Механизм восприятия звуков разной частоты.
В Внутреннее ухо содержит рецепторный аппарат двух анализаторов: вестибулярного (преддверие и полукружные каналы) и слухового, к которому относится улитка с кортиевым органом.
Костная полость внутреннего уха, содержащая большое число камер и проходов между ними, называется лабиринтом. Он состоит из двух частей: костного лабиринта и перепончатого лабиринта.Костный лабиринт – это ряд полостей, расположенных в плотной части височной кости; в нем различают три составляющие: полукружные каналы – один из источников нервных импульсов, отражающих положение тела в пространстве; преддверие; и улитку – орган слуха.
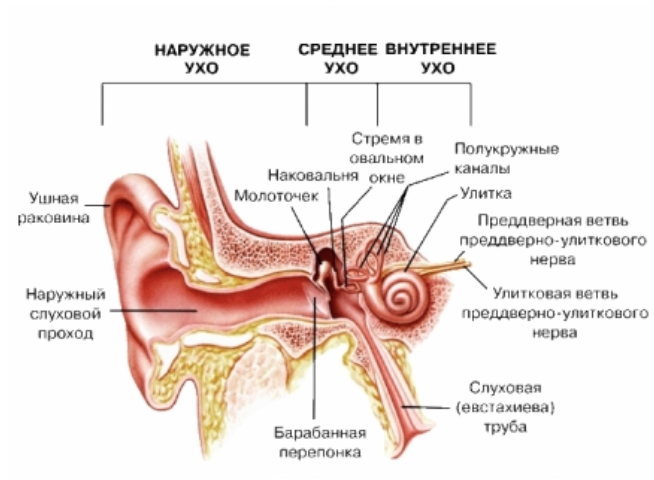
Строение уха
Перепончатый лабиринт заключен внутри костного лабиринта. Он наполнен жидкостью, эндолимфой, и окружен другой жидкостью – перилимфой, которая отделяет его от костного лабиринта. Перепончатый лабиринт, как и костный, состоит из трех основных частей. Первая соответствует по конфигурации трем полукружным каналам. Вторая делит костное преддверие на два отдела: маточку и мешочек. Удлиненная третья часть образует среднюю (улиточную) лестницу (спиральный канал), повторяющую изгибы улитки.
Полукружные каналы. Их всего шесть – по три в каждом ухе. Они имеют дугообразную форму и начинаются и кончаются в маточке. Три полукружных канала каждого уха расположены под прямыми углами друг к другу, один горизонтально, а два вертикально. Каждый канал имеет на одном конце расширение – ампулу. Шесть каналов расположены таким образом, что для каждого существует противолежащий ему канал в той же плоскости, но в другом ухе, однако их ампулы расположены на взаимнопротивоположных концах.
Улитка и кортиев орган. Название улитки определяется ее спирально извитой формой. Это костный канал, образующий два с половиной витка спирали и заполненный жидкостью. Завитки идут вокруг горизонтально лежащего стержня — веретена, вокруг которого наподобие винта закручена костная спиральная пластинка, пронизанная тонкими канальцами, где проходят волокна улитковой части преддверно-улиткового нерва — VIII пары черепно-мозговых нервов. Внутри, на одной стенке спирального канала по всей его длине расположен костный выступ. Две плоские мембраны идут от этого выступа к противоположной стенке так, что улитка по всей длине делится на три параллельных канала. Два наружных называются лестницей преддверия и барабанной лестницей, они сообщаются между собой у верхушки улитки. Центральный, т.н. спиральный, канал улитки, оканчивается слепо, а начало его сообщается с мешочком. Спиральный канал заполнен эндолимфой, лестница преддверия и барабанная лестница – перилимфой. Перилимфа имеет высокую концентрацию ионов натрия, тогда как эндолимфа – высокую концентрацию ионов калия. Важнейшей функцией эндолимфы, которая заряжена положительно по отношению к перилимфе, является создание на разделяющей их мембране электрического потенциала, обеспечивающего энергией процесс усиления входящих звуковых сигналов.
Лестница преддверия начинается в сферической полости – преддверии, лежащем в основании улитки. Один конец лестницы через овальное окно (окно преддверия) соприкасается с внутренней стенкой заполненной воздухом полости среднего уха. Барабанная лестница сообщается со средним ухом с помощью круглого окна (окна улитки). Жидкость
не может проходить через эти окна, так как овальное окно закрыто основанием стремени, а круглое – тонкой мембраной, отделяющей его от среднего уха. Спиральный канал улитки отделяется от барабанной лестницы т.н. основной (базилярной) мембраной, которая напоминает струнный инструмент в миниатюре. Она содержит ряд параллельных волокон различной длины и толщины, натянутых поперек спирального канала, причем волокна у основания спирального канала короткие и тонкие. Они постепенно удлиняются и утолщаются к концу улитки, как струны арфы. Мембрана покрыта рядами чувствительных, снабженных волосками клеток, составляющих т.н. кортиев орган, который выполняет высокоспециализированную функцию – превращает колебания основной мембраны в нервные импульсы. Волосковые клетки связаны с окончаниями нервных волокон, по выходе из кортиева органа образующих слуховой нерв (улитковую ветвь преддверно-улиткового нерва).
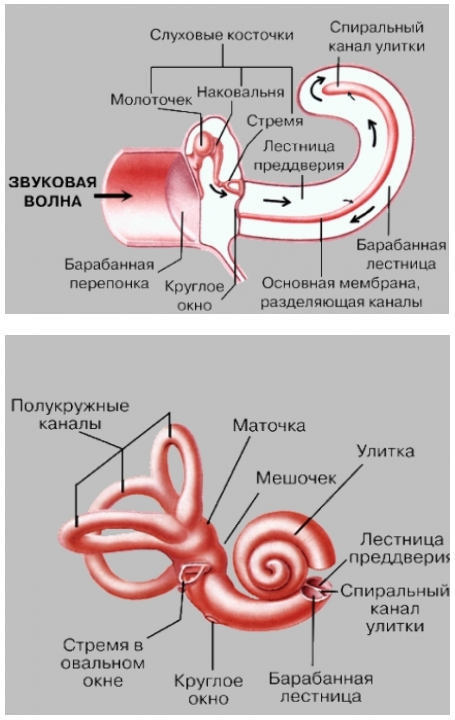
Улитка уха
Перепончатый улитковый лабиринт, или проток,имеет вид слепого преддверного выпячивания, находящегося в костной улитке и слепо заканчивающегося на ее верхушке. Он заполнен эндолимфой и представляет собой соединительно-тканный мешок длиной около35 мм. Улитковый проток разделяет костный спиральный канал на три части, занимая среднюю из них — средняя лестница (scala media), или улитковый ход, или улиточный канал. Верхняя часть — это лестница преддверия (scala vestibuli), или вестибулярная лестница, нижняя — барабанная, или тимпанальная, лестница (scala tympani). В них находится пери-лимфа. В области купола улитки обе лестницы сообщаются между собой через отверстие улитки (геликотрему). Барабанная лестница простирается до основания улитки, где она заканчивается у круглого окна улитки, закрытого вторичной барабанной перепонкой. Лестница преддверия сообщается с перилимфатическим пространством преддверия. Следует отметить, что перилимфа по своему составу напоминает плазму крови и цереброспинальную жидкость; в ней преобладает содержание натрия. Эндолимфа отличается от перилимфы более высокой (в 100 раз) концентрацией ионов калия и более низкой (в 10 раз) концентрацией ионов натрия; по своему химическому составу она напоминает внутриклеточную жидкость. По отношению к пери-лимфе она заряжена положительно.
Улитковый проток на поперечном разрезе имеет треугольную форму. Верхняя — преддверная стенка улиткового протока, обращенная к лестнице преддверия, образована тонкой преддверной (рейсснеровой) мембраной (membrana vestibularis), которая изнутри покрыта однослойным плоским эпителием, а снаружи — эндотелием. Между ними расположена тонкофибриллярная соединительная ткань. Наружная стенка срастается с надкостницей наружной стенки костной улитки и представлена спиральной связкой, которая имеется во всех завитках улитки. На связке расположена сосудистая полоска (stria vascularis), богатая капиллярами и покрытая кубическими клетками, которые продуцируют эндолимфу. Нижняя — барабанная стенка, обращенная к барабанной лестнице, устроена наиболее сложно. Она представлена базилярной мембраной, или пластинкой (lamina basilaris), на которой располагается спиральный, или кортиев орган, осуществляющий восприятие звуков. Плотная и упругая базиляр-ная пластинка, или основная мембрана, одним концом прикрепляется к спиральной костной пластинке, противоположным — к спиральной связке. Мембрана образована тонкими слабо натянутыми радиальными коллагеновыми волокнами (около 24 тыс.), длина которых возрастает от основания улитки к ее вершине — вблизи овального окна ширина базилярной мембраны составляет0,04 мм, а затем по направлению к вершине улитки, постепенно расширяясь, она достигает в конце0,5 мм(т.е. базилярная мембрана расширяется там, где улитка сужается). Волокна состоят из тонких анастомозирующих между собой фибрилл. Слабое натяжение волокон базилярной мембраны создает условия для их колебательных движений.
Собственно орган слуха — кортиев орган — находится в костной улитке.Кортиев орган — рецепторная часть слухового анализатора, расположенная внутри перепончатого лабиринта. В процессе эволюции возникает на основе структур боковых органов. Воспринимает колебания волокон, расположенных в канале внутреннего уха, и передаёт в слуховую зону коры больших полушарий, где и формируются звуковые сигналы. В Кортиевом органе начинается первичное формирование анализа звуковых сигналов.
Расположение.Кортиев орган располагается в спирально завитом костном канале внутреннего уха — улитковом ходе, заполненном эндолимфой и перилимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке. Кортиев орган представлен опорными, или поддерживающими, клетками, и рецепторными клетками, или фонорецепторами. Выделяют два типа опорных и два типа рецепторных клеток — наружные и внутренние.
Наружные опорные клетки лежат дальше от края спиральной костной пластинки, а внутренние — ближе к нему. Оба вида опорных клеток сходятся под острым углом друг к другу и образуют канал треугольной формы — внутренний (кортиев) туннель, заполненный эндо-лимфой, который проходит спирально вдоль всего корти-ева органа. В туннеле расположены безмиелиновые нервные волокна, идущие от нейронов спирального ганглия.
Фонорецепторы лежат на опорных клетках. Они представляют собой вторично-чувствующие рецепторы (механорецепторы), трансформирующие механические колебания в электрические потенциалы. Фонорецепторы (на основании их отношения к кортиеву туннелю) подразделяются на внутренние (колбообразной формы) и наружные (цилиндрической формы), которые отделены друг от друга кортиевыми дугами. Внутренние волосковые клетки располагаются в один ряд; их общее число по всей длине перепончатого канала достигает 3500. Наружные волосковые клетки располагаются в 3-4 ряда; их общее число достигает 12000-20000. Каждая волосковая клетка имеет удлиненную форму; один ее полюс приближен к основной мембране, второй находится в полости перепончатого канала улитки. На конце этого полюса есть волоски, или стереоцилии (до 100 в клетке). Волоски рецепторных клеток омываются эндолимфой и контактируют с покровной, или текториальной, мембраной (membrana tectoria), которая по всему ходу перепончатого канала расположена над волосковыми клетками. Эта мембрана имеет желеобразную консистенцию, один край которой прикрепляется к костной спиральной пластинке, а другой свободно оканчивается в полости улиткового протока чуть дальше наружных рецепторных клеток.
Все фонорецепторы, независимо от локализации, синаптически связаны с 32000 дендритов биполярных чувствительных клеток, находящихся в спиральном нервном ганглии улитки. Эти первыенейроны слухового пути, аксоны которых образуют улитковую (кохлеарную) часть VIII пары черепно-мозговых нервов; они передают сигналы на кохлеарные ядра продолговатого мозга. При этом сигналы от каждой внутренней волосковои клетки передаются на биполярные клетки одновременно по нескольким волокнам (вероятно, это повышает надежность передачи информации), в то время как сигналы от нескольких наружных волосковых клеток конвергируют на одном волокне. Поэтому около 95% волокон слухового нерва несет информацию в продолговатый мозг от внутренних волосковых клеток (хотя их количество не превышает 3500), а 5% волокон передают информацию от наружных волосковых клеток, число которых достигает 12000-20000. Эти данные подчеркивают огромную физиологическую значимость внутренних волосковых клеток в рецепции звуков.
К волосковым клеткам подходят и эфферентные волокна — аксоны нейронов верхней оливы. Волокна, приходящие к внутренним волосковым клеткам, оканчиваются не на самих этих клетках, а на афферентных волокнах. Предполагается, что они оказывают тормозное воздействие на передачу слухового сигнала, способствуя обострению частотного разрешения. Волокна, приходящие к наружным волосковым клеткам, воздействуют на них непосредственно и за счет изменения их длины, меняют их фоночувствительность. Таким образом, с помощью эфферентных оливо-кохлеарных волокон (волокон пучка Расмуссена) высшие акустические центры регулируют чувствительность фонорецепторов и поток афферентных импульсов от них к мозговым центрам.
Проведение звуковых колебаний в улитке.Восприятие звука осуществляется с участием фонорецепторов. Их возбуждение под влиянием звуковой волны приводит к генерации рецепторного потенциала, который вызывает возбуждение дендритов биполярногонейрона спирального ганглия. Но каким образом осуществляется кодирование частоты и силы звука? Это один из наиболее сложных вопросов физиологии слухового анализатора.
Современное представление о кодировании частоты и силы звука сводится к следующему. Звуковая волна, воздействуя на систему слуховых косточек среднего уха, приводит в колебательное движение мембрану овального окна преддверия, которая, прогибаясь, вызывает волнообразные перемещения перилимфы верхнего и нижнего каналов, которые постепенно затухают по направлению к вершине улитки. Поскольку все жидкости несжимаемы, колебания эти были бы невозможны, если бы не мембрана круглого окна, которая выпячивается при надавливании основания стремечка на овальное окно и принимает исходное положение при прекращении давления. Колебания перилимфы передаются на вестибулярную мембрану, а также на полость среднего канала, приводя в движение эндолимфу и базиляр-ную мембрану (вестибулярная мембрана очень тонкая, поэтому жидкость в верхнем и среднем каналах колеблется так, как будто оба канала едины). При действии на ухо звуков низкой частоты (до 1000 Гц) происходит смещение базилярной мембраны на всем ее протяжении от основания до верхушки улитки. При увеличении частоты звукового сигнала происходит перемещение укороченного по длине колеблющегося столба жидкости ближе к овальному окну, к наиболее жесткому и упругому участку базилярной мембраны. Деформируясь, базилярная мембрана смещает волоски волосковых клеток относительно текториальной мембраны. В результате такого смещения возникает электрический разряд волосковых клеток. Существует прямая зависимость между амплитудой смещения основной мембраны и количеством вовлекаемых в процесс возбуждения нейронов слуховой коры.
| Механизм проведения звуковых колебаний в улитке Звуковые волны улавливаются ушной раковиной и через слуховой канал направляются к барабанной перепонке. Колебания барабанной перепонки, через систему слуховых косточек, передаются посредством стремечка мембране овального окна, и через нее передаются лимфатической жидкости. На колебания жидкости отзываются (резонируют), в зависимости от частоты колебаний, только определенные волокна главной мембраны. Волосковые клетки Кортиева органа возбуждаются от прикосновения к ним волокон главной мембраны и по слуховому нерву передаются в мозгимпульсы, где и создается окончательное ощущение звука. http://www.braintools.ru/article/9775 |
Пути проведения звука. С функциональной точки зрения орган слуха (периферическая часть слухового анализатора) делится на две части:
1. звукопроводящий аппарат - наружное и среднее ухо, а также некоторые элементы (перилимфа и эндолимфа) внутреннего уха;
2. звуковоспринимающий аппарат - внутреннее ухо.
12. Синапсы, их классификация, строение. Механизм проведения возбуждения через возбуждающие и тормозные синапсы. Свойства синапсов.
Си́напс[1] (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Передача импульсов осуществляется химическим путём с помощью медиаторов или электрическим путём посредством прохождения ионов из одной клетки в другую.
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном. Однако сам Шеррингтон утверждал, что получил идею этого термина в разговоре от физиолога Майкла Фостера[2].

Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е —плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона.
Дата добавления: 2015-01-15; просмотров: 453; Мы поможем в написании вашей работы!; Нарушение авторских прав |