КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Поперечнополосатые мышечные ткани
Как показывает само название, особенность поперечнополосатой мышечной ткани — поперечная исчерченность, видимая даже при слабом увеличении светового микроскопа.
СКЕЛЕТНАЯ МЫШЕЧНАЯ ТКАНЬ
Характерной особенностью скелетных мышц является то, что они состоят не из клеток, а из крупных симпластических образований — мышечных волокон. По форме мышечное волокно напоминает цилиндр, достигающий в длину 12,5 см, eгo диаметр около 100 нм. Концы волокна закруглены или отдают несколько небольших отростков. В волокне различают оболочку (сарколемму), более сотни ядер и цитоплазму (саркоплазму), содержащую сократимое вещество. В саркоплазме мышечных волокон обнаруживается и ряд других структур: митохондрии, микросомы (рибосомы), трубочки и цистерны саркоплазматического ретикулума, различные вакуоли, глыбки гликогена и включения липидов, играющие роль запасных энергетических материалов, и т. д.
Сарколемма (sarcos — мясо, lemma — кожица, скорлупа) — очень тонкая оболочка, лучше всего обнаруживаемая в местах разрыва содержимого волокна. С помощью электронного микроскопа в ней удается различить два листка, разделенных светлым промежутком шириной 140—240 А.

Рисунок 3. Структура волокна скелетной мышцы (по Хассельбаху).
I- А-диск; II- I-диск; III- Н-зона; 1- Z-линия; 2- Т-система; 3- цитоплазматический ретикулум; 4- устье Т-системы; 5- гликоген; 6- митохондрии.
Наружный листок представляет собой базальную мембрану, к которой прикреплены коллапеновые фибриллы, располагающиеся в виде решетки; вследствие такого расположения фибрилл мышечное волокно может свободно увеличиваться в длину (при расслаблении) ив ширину (при сокращении).
Внутренний листок сарколеммы является плазмалеммой мышечного волокна. Его основная особенность — способность к распространению состояния возбуждения на все мышечное волокно. Деполяризация плазмалеммы, возникшая в одном мосте (в норме таким местом является двигательное нервное окончание), мгновенно волнообразно распространяется во всей толще мышечного волокна. Саркоплазма (цитоплазма) оказывается пронизанной системой поперечных трубок или Т-системой.
Ядра мышечного волокна оттеснены миофибриллами к самой периферии и находятся под сарколеммой. Иногда они лежат попарно или одно за другим и виде длинной цепочки, что указывает на их способность к амитотическому делению (амитозу). Форма ядер овальная; хроматин распылен мелкими зернами.
Саркоплазма окружает каждое ядро и заполняет промежутки между сократимым веществом. Ее количествосильно варьирует. В одних мышечных волокнах саркоплазмы относительно мало, зато много сократимого вещества, и такие волокна могут сокращаться сильно, но из-за невозможности быстрого восстановления затраченной энергии легко утомляются. Длительно работающие мышечные волокна, наоборот, богаты саркоплазмой и относительно бедны сократимым веществом. Саркоплазма содержит митохондрии, часто называемые здесь саркосомами, комплекс Гольджи и эндоплазматическую сеть.
Сократимое вещество имеет сложное строение, для лучшего понимания которого целесообразно рассмотреть схему строенияодной миофибриллы. Сократимое вещество и образованные им миофибриллы обусловливают поперечную исчерчонность мышечного волокна. Это вещество представлено белковыми нитями — более тонкими нитями актина и более толстыми — миозина, расположенными чередующимися участками. Участки миофибрилл с миозиновыми миофиламентами благодаря большей толщине последних будут в большей мере заполненными этими нитевидными структурами. Вещество, состоящее из параллельно расположенных частиц, оказывается оптически анизотропным, то есть с разной скоростью пропускает световые волны, колеблющиеся в разных направлениях. При рассматривании в обычный световой микроскоп такие участки будут более темными (оптическую анизотропию изучают с помощью поляризационного микроскопа). Описываемые участки миофибрилл называют анизотропными дисками — дисками А. В середине каждого диска А из-за утолщения миозиновых нитей во многих случаях образуется «срединная полоска» — полоска М, или мезофрагма (mesos — средний, phragma — перегородка). Полоска М расположена в середине сегмента миофибриллы.
Диски А чередуются с дисками, содержащими более тонкие активные миофиламенты. Они не обладают оптической анизотропией и называются поэтому изотропными дисками (дисками И). При рассматривании в обычный снеговой микроскоп диски И кажутся более светлыми. Диски И перегорожены полоской Z, или телофрагмой (telos — край). Возникает вопрос, почему полоски Z, тоже расположенные в середине диска, называют не срединными, а краевыми? Объясняется это сегментарностыо строения миофибрилл. Если подвергнуть мышечное волокно мацерации, оно все распадется на сегменты — сарком еры (meros — часть). Границами саркомера оказываются полоски Z, богатые мукополисахаридом и сравнительно легко расщепляющиеся при мацерации. Телофрагмы являются связующими участками между смежными сегментами, к ним с обеих сторон прикрепляются актиновые нити. Таким образом, в состав саркомера входят две телофрагмы, отграничивающие саркомер с обоих концов, к каждой из них прилегает по половине диска И. Среднюю часть саркомера занимает диск А, разделенный пополам полоской М.
Так саркомер выглядит только в условиях сильного расслабления мышечного волокна. В нормальном состоянии концы актиновых и миозиновых филаментов несколько заходят один за другой, образуя так называемую зону перекрытия, кажущуюся наиболее темной, так как в ней имеется больше всего миофиламентов (и активноых, и миозиновых).

Рисунок 4. Различные функциональные состояния саркомера.
I – расслабленная мышца; II – состояние слабого сокращения; III – сильное сокращение. И – изотропный и А – анизотропный диски4 Т – телофрагма; М – мезофрагма; ЗП – зона перекрытия;.В правой части показаны поперечные срезы через диски И, А и зону перекрытия.
Чем сильнее сокращение, тем глубже актиновые нити заходят промежутки между миозиновыми, зоны перекрытия расширяются за счет сужения полосок или зон Н. При полном сокращении мышечного волокна актиновые и миозиновые нити полностью зайдут одна за другую, весь саркомер превратится в общую зону перекрытия. Миофибриллы на всем их протяжении окажутся анизотропными, и диски И исчезнут. В зоне перекрытия между миозиновымн и актиновыми нитями возникают правильно расположенные связующие мостики. Очевидно, во время скольжения нитей мостики также меняют свое местоположение. Считают, что в области мостиков происходит распад молекул АТФ с освобождением энергии, необходимой для сокращения. Этот распад может происходить только в присутствии ионов кальция. Для понимания механизма сокращения необходимо знать, что в саркоплазме сильно развитая система тонкостенных трубочек и пистерн эндоплазматической (саркоплазматической) сети, синтезирующей гликоген и содержащей Са2+-ионы. Цистерны этой сети расположены между миофибриллами, то есть параллельно продольной оси волокна. В поперечном направлении через волокно проходят Т-трубки — впячивания внутреннего листка сарколеммы. Эти две системы цистерн и трубок взаимно перекрещиваются, вступая в тесный контакт в местах перекреста и обусловливая этим образование «триад» и «диад» мышечного волокна. Например, у лягушки Т-трубки проходят на уровне, полосок Z. На этом же уровне в каждом саркомере оканчиваются мешковидным расширением цистерны эндоплазматической сети.
Таким образом, Т-трубки, идущие по границе между двумя смежными саркомерами, контактируют с двух сторон с расширенными концами цистерн саркоплазматической сети обоих саркомеров. Такое сочетание трех трубчатых структур (Т-трубок и цистерн двух смежных саркомеров) называют триадой. В мышечных волокнах млекопитающих саркомеры длиннее, и через каждый из них проходят две Т-трубки на уровне обеих зон перекрытия. Понятно, что каждая Т-трубка может вступать контакт только с саркоплазматической сетью данного саркомера. Поэтому, в отличие от мышц лягушек, у млекопитающих образуются не триады, а диады.
Считают, что ионы кальция скапливаются в участках саркоплазматической сети в области триад или диад. Деполяризация плазмалеммы, вызванная возбуждением нерва, мгновенно распространяется по Т-системе вглубь мышечного волокна, врезультате этого увеличивается проницаемость мембраны саркоплазматической сети для Са2+-нонов. В присутствии этих ионов происходят распад молекул АТФ с освобождением энергии и сокращение волокна, восстановлением нормальной разности потенциалов на мембране Т-системы и плазмалемме ионы кальция возвращаются в саркоплазматическую сеть, мышечное волокно расслабляется.
Благодаря Т-трубкам и саркоплазматической сети все миофибриллы данного волокна удерживаются в определенном положении. Светлые и темные диски всех миофибрилл оказываются расположенными на одинаковых уровнях, и все волокно кажется поперечно исчерченным.

Рисунок 5. Мышечные волокна в поперечном разрезе: равномерное распределение миофибрилл; не равномерное распределение миофибрилл в виде полей Конгейма.
В одних случаях волокна сокращаются энергично, но быстро утомляются, в других наблюдается противоположная картина- Высокочастотные волокна сокращаются всего лишь на 4% своей длины, а низкочастотные — на 40%. Громадное разнообразие структурных отношении, связанное с физиологическими свойствами мышцы, с высотой организации и образом жизни животного, невозможно уложить в рамки простой схемы. На поперечном разрезе в одних волокнах миофибриллы распределены равномерно, в других они собираются пучками, имеющими в поперечном сечении вид многогранных, иногда причудливой формы нолей (поля Конгейма).
Даже в одной и той же мышце отмечают быстро и медленно сокращающиеся волокна. «Быстрые» волокна, в отличие от «медленных», имеют сильно развитую саркоплазматическую сеть, и поэтому миофибриллы четко отграничены. Митохондрий (саркосом) в них больше, саркомеры длиннее, полоски Z уже и яснее очерчены. Кроме того, быстрое волокно имеет только одно, но высоко-специализированное двигательное нервное окончание — моторную бляшку, медленные волокна снабжены несколькими примитивными двигательными окончаниями.
Концы мышечных волокон прочно прикрепляются к сухожилию или к сухожильным прослойкам, расположенным внутри мышцы. В этом месте мышечное волокно образует пальцевидные выступы, вдающиеся в сухожилие. Упоминавшиеся выше соединительнотканые фибриллы, одевающие снаружи сарколемму, также участвуют и образовании связи с сухожилием. Эта связь настолько прочна, что не нарушается даже при растяжении, способном разорвать мышцу или сухожилие.
Что касается связи отдельных мышечных волокон друг с другом, то она, как и в гладкой мускулатуре, осуществляется соединительной тканью. Снаружи вся мышца окружена соединительнотканной оболочкой (перимизий), от которой внутрь отходят более тонкие прослойки этой же ткани, одевающей каждое мышечное волокно оболочкой (эндомизий. Мышца, рассматриваемая как орган, вовсе не простая совокупность мышечных волокон; это — сложная система мышечных и соединительнотканых элементов, содержащая множество кровеносных сосудов и нервов. Посредством сухожилий мышца соединяется с частями скелета, образуя вместе с последними орган движения.
 Сердечная мышца (миокард)
Сердечная мышца (миокард)
Сердечная мышца, не утомляясь, ритмично работает в течение всей жизни животного. Предсердия и желудочки сокращаются неодновременно, и в то же время их движения строго согласованы.
Последовательность сокращений предсердий и желудочков, сохраняющаяся даже и изолированном сердце, во многом обусловлена тем, что эти отделы связаны между собой мышечными тяжами особого строения, известными под названием «проводящая мускулатура сердца». Таким образом, различают два вида сердечной мускулатуры — основную массу мышечной ткани сердца, обеспечивающую сокращение этого органа и называемую поэтому рабочей мускулатурой сердца, и проводящую мускулатуру, благодаря которой достигается согласованность сокращении предсердий и желудочков. Прирассматривании в световой микроскоп сердечная мышца имеет вид симпласта, состоящего из одетых сарколеммой анастомозирующих между собой мышечных волокон, между которыми располагается богатая сосудами и нервами рыхлая соединительная ткань. Волокна содержат очень много саркоплазмы, богатой гликогеном. Вследствие обилия саркоплазмы находящиеся в ней макроэргические вещества (вещества, освобождающие при своем разложении большое количество энергии) вполне обеспечивают беспрерывность деятельности сердца.
С этой особенностью строения сердечной мышцы связано и своеобразное расположение ядер. Они лежат не под сарколеммой, а в центре мышечного волокна, тогда как миофибриллы, концентрирующиесяпучками,проходят ближе к периферии, что хорошо видно на поперечном срезе. При относительно небольшом числе миофибрилл продольная исчерченность мышечной ткани выражена довольно резко, а поперечная сравнительно слабо.

Рисунок 6. Проводящая мускулатура сердца овцы.
а – проводящая мускулатура; в – рабочая мускулатура.
Никакого симпласта сердечная мышца не образует. Все мышечные волокна состоят из отдельных одноядерных мышечных клеток, располагающихся цепочкой одна за другой. Взаимное соединение клеток осуществляется специальным приспособлением, очень сходным с десмосомами эпителия. Границы между клетками образованы внутренним слоем сарколеммы, то есть плазмалеммой. Эти границы топографически совпадают с полосками Z. В одних случаях граница может идти прямо и полностью совпадать с одной полоской Z, вдругих она идет ступенчато, переходя с одной полоски Z на следующую. Вместах прикрепления миофибрилл межклеточные границы утолщены. Десмосомы образуются там, где к поверхностной мембране прикрепляются пучки тонофиламентов. Сходство с десмосомами увеличивается еще и тем, что все эти образования обычно богаты мукополисахаридами.
С возрастом одновременно с увеличением количества миофибрилл всердечной мышце происходит иутолщение клеточных границ. Это особенно сильно выражено в мускулатуре желудочков, в частности у крупного рогатого скота и лошадей, а также у человека. Утолщение границ может быть настолько сильным, что они становятся видимыми в световой микроскоп.
Однако контактов между Т-трубкамн и саркоплазматической сетью в сердечной мышце заметно меньше. Поэтому для сокращения последней требуется приток ионов кальция извне. Известно, что введение в кровь соли кальция усиливает работу сердца.
Новейшие данные о клеточном строении сердечной мышцы подтверждаются многими факторами. В культурах этой ткани две клетки, отделенные друг от друга вставочной полоской, сокращаются с различной скоростью; многие патологические изменения (набухание и пр.) могут развиваться только в одной клетке, не переходя в соседние.
Проводящая мускулатура сердца у животных разных видов различного строения; она особенно сильно выражена у животных с редким пульсом, в частности у жвачных. Тяжиэтой мускулатуры отличаются светлой окраской и неправильными очертаниями, по ходу их нередко встречаются расширенные участки, в которых залегают ядра (часто попарно). Миофибрилл мало и идут они не параллельно, а перекрещиваясь под острым углом. Описываемая разновидность сердечной практически лишена Т-системы, так как к активному сокращению она не способна.
Проводящая мускулатура, как, впрочем, и вся сердечная мышца, особенно богата нервными волокнами — двигательными и чувствительными нервами. По характеру связи с нервной системой сердечная мышца резко отличается от скелетной мускулатуры более приближаясь в этом отношении к гладкой мышечной ткани. Проводящая мускулатура содержит еще больше саркоплазмы. По сравнению с рабочей мускулатурой она обладает еще большим количеством гликогена, но бедна саркосомами (митохондриями). Так как с последними связана ферментативная активность живого вещества, этой бедностью саркосомами и объясняется слабая выраженность окислительных процессов. Проводящая мускулатура потребляет мало кислорода, довольствуясь относительно примитивным бескислородным расщеплением гликогена. Гликоген проводящей мускулатуры отличается от гликогена других органов большей устойчивостью к кислотам и меньшей растворимостью в воде.

Рисунок 7. Мышца сердца.
1 – ядро; 2 – прослойка соединительной ткани; 3 – вставочные полоски.
Наблюдения над заживлением повреждений миокарда показывают, что в большинстве случаев отмирающий участок мышцы замещается соединительной тканью, гораздо быстрее регенерирующей, чем мышечная ткань. Значение этой приспособительной реакции вполне понятно, сердце — настолько важный для животного орган, что всякая задержка с закрытием образовавшегося дефекта может привести к смертельному исходу.
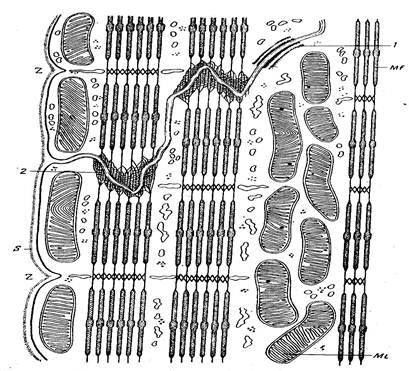
Рисунок 8. Схема строение сердечной мышцы мыши в области вставочной полоски.
S – сарколемма; Mi – митохондрия; Mf – микрофиламенты; 1 – зона уплотнения на плазмалемме; 2 – окончание миофибриллы на поазмалемме; Z - телофрагмы.
Дата добавления: 2015-02-10; просмотров: 869; Мы поможем в написании вашей работы!; Нарушение авторских прав |