КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Небелковые азотистые экстрактивные вещества
В скелетных мышцах содержится ряд важных азотистых экстрактивных веществ: адениновые нуклеотиды (АТФ, АДФ и АМФ), нуклеотиды неаденинового ряда, креатинфосфат, креатин, креатинин, карнозин, анзерин, свободные аминокислоты и др. Концентрация адениновых нуклеотидов в скелетной мускулатуре кролика (в мкмоль на 1 г сырой массы ткани) составляет: АТФ —4,43, АДФ —0,81; АМФ —0,93. Количество нуклеотидов неаденинового ряда (ГТФ, УТФ, ЦТФ и др.) в мышечной ткани по сравнению с концентрацией адениновых нуклеотидов очень мало.
На долю азота креатина и креатинфосфата приходится до 60% небелкового азота мыши [Фердман Д. Л., 1966]. Креатин-фосфат и креатин относятся к тем азотистым экстрактивным веществам мышц, которые участвуют в химических процессах, связанных с мышечным сокращением.
Напомним, что синтез креатина в основном происходит в печени, откуда он с током крови поступает в мышечную ткань. Здесь креатин, фосфорилируясь, превращается в креатинфосфат. В синтезе креатина участвуют три аминокислоты: аргинин, глицин и метионин. На схеме изображены основные этапы образования креатина и креатинфосфата.
К числу азотистых веществ мышечной ткани принадлежат и имидазолсодержащие дипептиды — карнозин и анзерин. Карнозин был открыт В. С. Гулевичем в 1900 г. Метилированное производное карнозина — анзерин был обнаружен в мышечной ткани несколько позже.
Карнозин и анзерин — специфические азотистые вещества скелетной мускулатуры позвоночных — увеличивают амплитуду мышечного сокращения, предварительно сниженную утомлением. Принято считать, что имидазолсодержащие дипептиды не влияют непосредственно на сократительный аппарат, но; увеличивают эффективность работы ионных насосов мышечной клетки (Северин С. Е.).
Из свободных аминокислот в мышцах наиболее высокая концентрация глутаминовой кислоты (до 1,2 г/кг) и ее амида — глутамина (0,8—1,0 г/кг). В состав различных клеточных мембран мышечной ткани входит ряд фосфатидов:фосфатидилхолив, фосфатидилэтаноламин, фосфатиднлсерив и др. Кроме того фосфатиды принимают участие в обменных процессах, в частности, в качестве субстратов тканевого дыхания. Другие азотсодержащие вещества: мочевина, мочевая кислота, аденнн гуанин, ксантин и гипоксантин — встречаются в мышечной ткани в небольшом количестве и, как правило, являются либо промежуточными, либо конечными продуктами азотистого обмена.
Безазотистые вещества мышц.Одним из основных представителей безазотистых органических веществ мышечной ткани является гликоген; его концентрация колеблется от 0,3% до 2% и выше. На долю других представителей углеводов приходятся десятые и сотые доли процента. В мышцах находят лишь следы свободной глюкозы и очень мало гексозофосфатов. В процессе метаболизма глюкозы, а также аминокислот в мышечной ткани образуются молочная, пировиноградная кислоты и много других карбоновых кислот. В мышечной ткани обнаруживаются также в том или ином количестве нейтральные жиры и холестерин.
Состав неорганических солей в мышцах разнообразен. Среди катионов наибольшую концентрацию имеют калий и натрий. Калий сосредоточен главным образом внутри мышечных волокон, а натрий — преимущественно в межклеточном веществе. Значительно меньше в мышцах магния, кальция и железа. В мышечной ткани содержится ряд микроэлементов: кобальт, алюминий, никель, бор, цинк и др.
Функциональная биохимия мышц.Мышечный аппарат человека и животных характеризуется полифункциональностью. Однако основной функцией мышц является осуществление двигательного акта, т.е. сокращение и расслабление. При сокращении мышц осуществляется работа, связанная с превращением химической энергии в механическую.
Источники энергии мышечной деятельности.В настоящее время принято считать, что процессом, непосредственно связанным с работающим механизмом поперечнополосатого мышечного волокна, является распад АТФ с образованием АДФ и неорганического фосфата. Возникает вопрос каким образом мышечная клетка может обеспечить свой сократительный аппарат достаточным количеством энергии в форме АТФ? Точнее: каким образом в процессе мышечной деятельности происходит непрерывный ресинтез этого макроэрга?
Прежде всего ресинтез АТФ обеспечивается трансфосфорилированием АДФ с креатинфосфатом. Данная реакция катализируется ферментом креатинкиназой:
Креатинфосфат + АДФ « креатин + АТФ
Креатинкиназа
Креатинкиназный путь ресинтеза АТФ является чрезвычайно быстрым и максимально эффективным (за счет каждой молекулы креатинфосфата образуется молекула АТФ). Именно поэтому долгое время не удавалось установить снижение концентрации АТФ и соответственно повышение концентрации АДФ даже при достаточно продолжительном тетанусе.
Некоторое количество АТФ может ресинтезироваться в ходе аденилаткиназной (миокиназной) реакции:

 2 АДФ АТФ + АМФ
2 АДФ АТФ + АМФ
аденидаткиназа
Запасы креатинфосфата в мышце невелики, а доступность анергии креатинфосфата имеет ценность для работающей мышцы, только в том случае, если расход его постоянно возмещается синтезом АТФ в процессе метаболизма. Для любой ткани, в том числе и мышечной, известно два фундаментальных биохимических процесса, в ходе которых регенерируются богатые энергией фосфорные соединения. Один из этих процессов — гликолиз, другой — тканевое дыхание. Наиболее важным и эффективным из них является тканевое дыхание. При достаточном снабжении кислородом мышца, несмотря на анаэробный механизм сокращения, в конечном итоге работает за счет энергии, образующейся при окислении (в цикле Кребса) как продуктов распада углеводов, так и ряда других субстратов тканевого дыхания, в частности жирных кислот, а также ацетата.
В последнее время появились данные, доказывающие, что креатинфосфат в мышечной ткани (в частности, в сердечной мышце) способен выполнять не только роль как бы депо легкомобилизируемых макроэргических фосфатных групп, но и играть также роль транспортной формы макроэргических фосфатных связей, образующихся в процессе тканевого дыхания и связанного с ним окислительного фосфорилирования.
При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробного метаболизма. Однако при больших нагрузках, когда возможность снабжения кислородом отстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщепления гликогена или глюкозы с образованием молочной кислоты увеличивается в сотни раз. Соответственно содержание молочной кислоты в мышечной ткани может повышаться до 1—1,2 г/кг и выше. Последняя с током крови в значительном количестве поступает в печень, где ресиитезируется в глюкозу и гликоген за счет энергии окислительных процессов. Перечисленные механизмы ресинтеза АТФ при мышечной деятельности включаются в строго определенной последовательности.
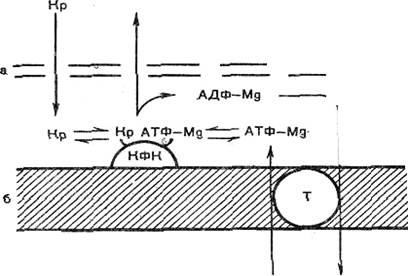
Рисунок 11. Схема переноса энергии из митохондрий в цитоплазму клетки миокарда (В. Н. Сакс и др.).
Наиболее экстренным является креатинкиназный механизм, илишь примерно через 20 с максимально интенсивной работы начинается усиление гликолиза, интенсивность которого достигает максимума через 40—80 с. При работе более длительной, а следовательно, и менее интенсивной,- все большее значение приобретает аэробный путь ресинтеза АТФ.
Содержание АТФ и креатинфосфата в сердечной мышие ниже, чем в скелетной мускулатуре, а расход АТФ велик, поэтому ресинтез АТФ в миокарде должен проходить намного интенсивнее, чем в скелетной мускулатуре. Для сердечной мышцы теплокровных животных и человека основным путем образования образования богатых энергией фосфорных соединений является путь окислительного фосфорилирования, связанный с поглощением кислорода. Регенерация АТФ в процессе анаэробного расщепления углеводов (гликолиз) в сердце человека практического значения не имеет. Именно поэтому сердечная мышца очень чувствительна к недостатку кислорода. Характерной особенностью обмена веществ сердечной Мышцы по сравнению со скелетной мускулатурой является также то, что аэробное окисление веществ неуглеводной природы при работе сердечной мышцы имеет большее значение, чем при сокращении скелетной мышцы. Только 30—35% кислорода, поглощаемого сердцем в норме, расходуется на окисление углеводов и продуктов их превращения. Главным субстратом дыхания в сердечной мышце являются жирные кислоты. Окисление неуглеводных веществ обеспечивает около 65—70% потребности миокарда в энергии. Из свободных жирных кислот в сердечной мышце особенно легко подвергается окислению олеиновая кислота.
Дата добавления: 2015-02-10; просмотров: 504; Мы поможем в написании вашей работы!; Нарушение авторских прав |