КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Рыхлая неоформленная соединительная ткань
Эта ткань очень распространена в организме. Она сопровождает кровеносные сосуды, поэтому входит в состав почти всех органов; она же заполняет и промежутки между многими органами. Общеизвестным примером рыхлой неоформленнойсоединительной ткани служит подкожная клетчатка.
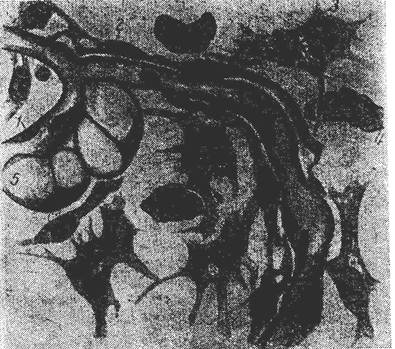
Рисунок 14. Рыхлая неоформленная ткань подкожной клетчатки кролика (по Г. Ясвоину):
1- эндотелий; 2- адвентициальная (камбиальная) клетка; 3- фибробласт; 4- гистиоцит; 5- жировая клетка.
В состав рыхлой соединительной ткани входят клетки и межклеточное вещество. Последнее состоит из аморфного вещества и волокон. Клеточные элементыдовольно разнообразны; среди них есть и собственные клетки соединительной ткани, и клетки кропи, эмигрировавшие из сосудов. В сложности состава нетрудно убедиться из схемы.
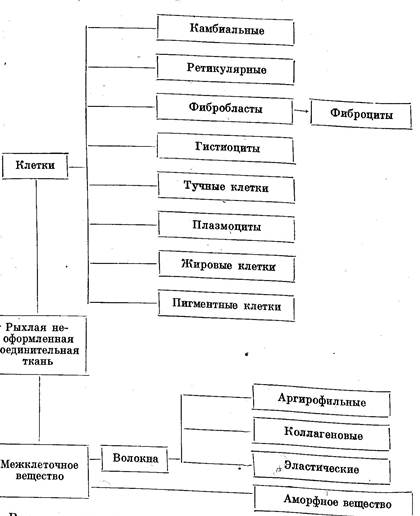
Малодифференцированные клетки соединительном ткани — камбиальные (стволовые) и ретикулярные. Степень дифференциации клеток соединительной ткани у животных разныхвидов и даже у одной и той же особи может быть различной. Наименее дифференцированные камбиальные клетки располагаются главным образом вокруг кровеносных капилляров. Они во многом напоминают мезенхимные элементы и представляют собой неправильной формы звездчатые клетки, часто с длинными отростками. Цитоплазма их окрашивается интенсивно и неравномерно; ядро небольшое, богатое хроматином. Эти клетки известны под названиями: камбиальные, адвентициальные (odventicus — пришлый), или мезенхимные (последнее название неудачно, так как у взрослых животных мезенхимы не бывает).

Рисунок 15. Рыхлая соединительная ткань: А – общий вид; Б – схема эластической сети; В - схема пучка эластических волокон; Г – снимок с препарата; 1 – фибробласт; 2 – гистиоцит; 3 – лейкоцит; 4 =- пучок колагеновых волокон; 5 – сеть эластических волокон.
Фибробласты — производные камбиальных клеток, обнаруживающие признаки более высокой дифференциации. Тело клетки уплощено и при рассматривании с поверхности имеет вид неправильной формы пластинки.
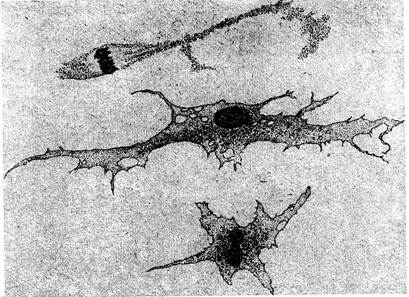
Рисунок 16. Фибробласты крупного рогатого скота (с препарата А.И. Тельцова): вверху – фибробласт, делящийся митозом; внизу – амитотическое деление ядра.
В поперечном разрезе фибробласт кажется веретенообразным, так как его центральная часть, содержащая ядро и эндоплазму, значительно утолщена. Довольно крупное овальное или округлое ядро окрашивается слабо, содержит равномерно распыленный хроматин и 2—3 отчетливо видимых ядрышка. Эндоплазма окрашивается интенсивно: в ней есть органоиды. Эктоплазма гомогенна и слабо окрашивается, вследствие чего контуры клетки нерезко очерчены и как бы переходят постепенно в аморфное межклеточное вещество. У взрослых животных с сильно развитой соединительной тканью эти клетки сохраняют лишь небольшое количество цитоплазмы; такие высокодифференцированные клетки обычно называют фиброцитами.
При электронной микроскопии в поверхностных слоях цитоплазмы нередко обнаруживаются уплощенные пузырьки цито-плазматической сети с многочисленными плотными гранулами на их стенке. Появление этих структур связано с образованием волокон соединительной ткани (см. ниже).
Гистиоциты встречаются примерно в таком же количестве, в каком и клетки фибробластического ряда. В отличие от фибробластов гистиоцит не имеет тесной связи с межклеточным веществом; границы клетки резко очерчены. Ядро относительно богато хроматином в виде крупных глыбок. Цитоплазма отчетливо контурирована и, кроме органоидов, часто содержит вакуоли и включения, свидетельствующие об активном участии этих клеток в обмене веществ и об их фагоцитарной деятельности (рис. 156). Если ввести в соединительную ткань окрашенные коллоидальные вещества, последние через короткий промежуток времени окажутся захваченными гистиоцитами. При воспалении эти клетки приходят в состояние раздражения, увеличиваются в размерах, обнаруживают способность к активному передвижению и превращаются в полибласты. Следовательно, по биологическим свойствам гистиоциты значительно отличаются от клеток фибробластического ряда. Их развитие протекает по иному пути. Фибробласты имеют непосредственное отношение к процессам образования межклеточного вещества, тогда как гистиоциты выполняют трофическую и защитную роль и этим значительно сближаются с ретикулярными клетками. Необходимо иметь в виду, что гистиоциты нередко описываются в литературе под другими названиями (блуждающие клетки в покое, клазматоциты, нефрофагоциты).
При электронной микроскопии на поверхности гистиоцитов видны длинные пластинчатые отростки. Фагоцитируемая клеткой частица охватывается этими отростками, затем отростки сливаются верхушками друг с другом, и захваченная частица оказывается включенной в цитоплазму и переваривается.
Отростки гистиоцитов могут иметь и иное значение. В местах, где гистиоцитов (или других макрофагов) много и они прилегают друг к другу, отростки одной клетки проникают между отростками соседней клетки.
При проникновении в соединительную ткань крупных чужеродных частиц макрофаги могут сливаться в одну гигантскую клетку инородного тела. Их слияние удается проследить в тканевых культурах. Впрочем, не исключена возможность образования гигантской клетки и путем многократного амитотического деления ядра одного макрофага.
Все остальные клетки соединительной ткани являются производными ретикулярных клеток, фибробластов и гистиоцитов или же эмигрируют из кровеносных сосудов.
Жировые клетки располагаются главным образом вблизи кровеносных сосудов, а местами формируют мощные отложения жировой ткани. Они являются, вероятнее всего, производными камбиальных и ретикулярных элементов, а также гистиоцитов, то есть клеток, активно участвующих в промежуточном обмене веществ и накапливающих в своей цитоплазме жиры. Жир появляется в них сначала в виде мелких капель, рассеянных в цитоплазме. Затем они сливаются в одну большую каплю, настолько крупную, что она заполняет почти все тело клетки. Цитоплазма и ядро при этом оттесняются к периферии, и жировая клетка приобретает своеобразную и типичную для нее форму, напоминающую на поперечном разрезе перстень (рис. 157). При применении специальных красителей (судан III и др.) жиры окрашиваются очень избирательно (по оттенку окраски можно судить о химическом составе жиров). На обычных препаратах, обработанных спиртом, жир растворяется и жировые клетки кажутся пустыми.
При электронной микроскопии бросаются в глаза почти полное отсутствие в цитоплазме эндоплазматической сети и слабое развитие зоны Гольджи. Эта особенность вполне понятна, так как упомянутые органоиды связаны преимущественно с синтезом белков, в жировых клетках почти отсутствующим. Впрочем, в буром жире эти органоиды развиты значительно сильнее.
Изредка слишком энергичное накапливание жира или какая-либо другая причина приводят к разрыву цитоплазмы жировой клетки. В этих случаях капля жира оказывается лежащей в межклеточном пространстве и окружена слоем плоских клеток (чаще всего фибробластов).
Тучные клетки(клетки Эрлиха, или гепариноциты) встречаются в различных частях тела, скапливаясь иногда в довольно большом числе. Количество их, возможно, связано с возрастом (замечено, например, что в мышцах крупного рогатого скота число тучных клеток с возрастом сильно увеличивается). Это — довольно крупные, большей частью неправильно-округлые клетки, характерным признаком которых служит наличие в цитоплазме многочисленных крупных базофильных зерен.
Как и у базофилов крови, эта зернистость растворима в воде и окрашивается некоторыми красителями (например, толуидиновым синим) метахроматически, то есть с изменением цвета красителя.
Между тучными клетками и базофилами наблюдается определенная зависимость: у животных, не имеющих в крови базофилов (крысы), в соединительной ткани находят много тучных клеток; у кролика базофилов в крови больше, чем у других млекопитающих, а тучных клеток в соединительной ткани очень мало.
В последнее время удалось доказать, что тучные клетки выделяют вещество типа гепарина. Гепарин — химическое соединение (мукополисахарид), образующееся в печени и в некоторых других органах, препятствующее свертыванию крови в организме. Известны опухоли, состоящие из сильно размножающихся тучных клеток. У страдающих этим заболеванием животных резко понижена свертываемость крови, ввиду чего они склонны к кровотечениям. Вытяжка, приготовленная из такой опухоли, содержит в 50 раз больше гепарина, чем вытяжка из нормальной печени. Добавление ее к извлеченной из организма крови резко замедляет свертываемость последней.
Известно еще одно важное свойство тучных клеток. Подобно базофилам крови, они содержат гистамин — чрезвычайно активное вещество, резко расширяющее кровеносные сосуды и этим снижающее кровяное давление, увеличивающее проницаемость капилляров, возбуждающее гладкие мышцы и др. Гистамин постоянно образуется в организме из аминокислоты гистидина и накапливается в гранулах базофилов и тучных клеток. При введении в организм препарата, разрушающего эти клетки, происходит немедленное освобождение большого количества гистамина, наступает опасное для жизни шоковое состояние. Подобное явление нередко возникает под влиянием чужеродных белков. Можно предполагать, что тучные клетки накапливают гистамин и разрушают его с помощью фермента гистаминазы.
В охлаждаемой конечности животного число тучных клеток уменьшается, так как многие из них распадаются, освобождая гистамин. Последний быстро удаляется из организма почками. При длительном содержании животных на холоде у них развивается устойчивость к гистамину. Возможно, что при этом меняются и свойства оставшихся тучных клеток.
Плазматические клетки более или менее округлой формы. Ядро лежит эксцентрично и содержит крупные глыбки хроматина. Цитоплазма, особенно в периферическом слое, резко базофильна, что объясняется обилием гранул РНК. Кроме гранул, электронная микроскопия выявляет в цитоплазме исключительно сильно развитую эндоплазматическую сеть из плоских цистерн. Сильное развитие этого органоида в сочетании с гранулами РНК свидетельствует об интенсивном синтезе белка. Установлено, что плазматические клетки выделяют белок γ-глобулин, имеющий непосредственное отношение к иммунитету (невосприимчивости к заразным заболеваниям). На проникновение чужеродного белка организм отвечает усиленным образованием γ-глобулина, синтезируемого плазматическими клетками (плазмоцитами). Высокая концентрация γ-глобулина часто приводит к его отложению в цитоплазме плазматических клеток в виде резко эозинофильных гранул и кристаллов, называемых тельцами Расела (часто их называют тельцами Русселя в соответствии с неправильной транскрипцией имени этого исследователя).
Таким образом, плазмоциты — типичные иммунокомпетентные клетки. При описании лимфоцитов говорилось, что при столкновении с антигеном лимфоциты превращаются в иммунокомпетентные клетки, часто называемые пиронинофильными, то есть окрашивающимися пиронином — красителем, выявляющим РНК. Эти Клетки снабжены сильно развитой гранулярной эндоплазматической сетью и вырабатывают защитные антитела, обычно еще примитивные и имеющие местное значение. Таковы, например, «клетки отторжения трансплантата», доставляющие немало неприятностей хирургам.
Пигментные клетки в нормальной соединительной ткани млекопитающих не встречаются. Пигментные клетки оболочек мозга и органа зрения, вероятно, не имеют прямого отношения к соединительной ткани. У низших позвоночных животных соединительнотканые пигментные клетки — хронатофоры- обнаруживают постоянно в виде неправильной формы отростчатых клеток. Их цитоплазма содержит пигмент. Под регулирующим влиянием нервной система хроматофоры могут активно сокращаться, вследствие чего изменяется цвет кожного покрова, что делает животное менее заметным.
Межклеточное вещество соединительной ткани. Эта составная часть соединительной ткани представляет собой своеобразную форму живого вещества, выполняющего важную роль. В нем, кроме аморфного вещества, имеются волокна — коллагеновые, эластические и аргирофильные (преколлагеновые). Как на волокнах, так и в аморфном веществе залегает много чувствительных нервных окончаний, непрерывно посылающих в центральную нервную систему сигналы о состоянии натяжения этих структур. Межклеточное вещество отнюдь не мертво, в нем происходит обмен веществ, осуществляется перестройка волокон в соответствии с изменением направления действия механических факторов.
Аморфное вещество тесно связано с фибробластами (фиброцитами) и образуется из веществ, вырабатываемых этими клетками. Оно появляется на самих ранних этапах развития соединительной ткани и первоначально состоит преимущественно из мукополисахаридов. Основные мукополисахариды — гепарин, гиалуроновая и хондроитинсерная кислоты. Гепарин содержится в гранулах тучных клеток и базофилов, гиалуроновая кислота — в молодом, еще мягком аморфном веществе рыхлой соединительной ткани, а хоидроитинсерная кислота — в плотной опорной (хрящевой) ткани. О наличии мукополисахаридов можно судить по способности данного вещества вызывать деполимеризацию молекул красителя и этим наменять их цвет. В частности, мукополисахариды окрашиваются толуидиновым синим не в синий, а в фиолетовый цвет. Вязкое аморфное вещество заполняет все промежутки между клетками и волокнами. Оно может связывать значительные количества воды; в нем происходит оживленный обмен веществ, в процессе которого отмешиваются волокнистые белки. В местах наибольшей активности аморфного вещества (стенка венечных артерий сердца, восходящая аорта) легче всего возникают и различные нарушения обмена, при которых вместо волокнистых белков начинают откладываться жироподобные вещества или минеральные соли. Так возникают тяжелые и опасные для организма изменения кровеносных сосудов.
Будучи бесструктурным и прозрачным, аморфное вещество не обнаруживается при световой .микроскопии. Его можно увидеть лишь вызвав отек ткани, при которой накопившаяся тканевая жидкость расщепляет аморфное вещество на тончайшие пластинки.
По мере развития соединительнотканных волокон количество аморфного вещества уменьшается, оно утрачивает мукополисахариды, а вместе с этим и прозрачность.
Коллагеновые волокна состоит из волокнистого белка — коллагена (colla — клей, gennao — я произвожу), получившего название из-за свойства прекращаться при длительном варении к животный клей желатин. Это — довольно толстые волокна, обычно не анастомозирующие между собой, идущие или в разных направлениях (рыхлая соединительная ткань), или параллельно друг другу в направлении действия сил, стремящихся растянуть данную ткань (плотная оформленная ткань). Они имеют продольную исчерченность, так как состоят из тонких коллагеновых фибрилл. Коллагеновое волокно представляет собой пучок фибрилл совершенно одинаковой толщины, погруженных в межфибриллярное цементирующее вещество. Благодаря правильному продольному расположению образующих фибриллы частиц коллагеновые волокна обнаруживают при исследовании в поляризационном микроскопе типичную оптическую анизотропию. Окрашиваются они как кислыми, так и основными красителями.

Рисунок 17. схема строения коллагенового волокна.
А – спиральная структура макромолекулы коллагена (по Ричу): мелкие светлые кружки – глицин, крупные светлые кружки – пролин, заштрихованные кружки – гидроксипролин.
Б – схема строения коллагеновых волокон (по Ричу): 1 – пучок фибрилл; 2 – фибрилла; 3 – протофибрилла; 4 - молекула коллагена.
Коллагеновые волокна очень прочны и почти не растяжимы, образованные ими сухожилия выдерживают нагрузку до 0 кг на 1 мм2 сечения. Помимо опорной функции, они выполняют роль своеобразного фильтра, так как могут адсорбировать на поверхности различные вещества.
При электронной микроскопии обнаруживается типичная для коллагеновых фибрилл поперечная исчерченность (рис. 17), свидетельствующая о периодичности их химического строения с длиной периода 610 А.
Коллагеновые фибриллы, в свою очередь, состоят из более тонких протофибрилл (филаментов), образованных молекулами коллагена. Каждый период, имеющий в длину 640 А, состоит из двух зон, светлой и темной. Светлая зона несколько длиннее темной, но для простоты можно принять, что каждая зона соответствует половине периода. Молекулы коллагена при полимеризации располагаются друг за другом в виде цепочки, но между двумя молекулами остается разрыв, соответствующий темному полупериоду (рис. 17). При «сшивании» своими боковыми поверхностями молекулы коллагена располагаются ступенчато, то есть молекулы одного ряда смещаются по отношению к молекулам другого ряда на половину периода (на темную половину). Благодаря этому создается впечатление поперечной исчерченности всей фибриллы.
Рентгеновский структурный анализ, позволяющий определить расстояния между отдельными группами атомов, показывает, что каждая молекула состоит из трех совершенно одинаковых полипептидных нитей. На рисунке 17 представлена модель молекулы тропоколлагена (примитивного коллагена). В каждой из трех спирально скрученных полипептидных нитей видно правильное чередование трех аминокислот глицина, пролина и гидроксипролина (другие аминокислоты встречающиеся в меньших количествах, на схеме не показаны).
Молекулярный вес каждой полипептидной нити 120 000, а всей макромолекулы - 360 000. Все три нити соединены между собой боковыми, преимущественно водородными, связями. Под действием мочевины, разрушающей эти связи, молекулы распадаются на отдельные полипептидные нити.
Спиральное скручивание может наблюдаться не только у полипептидных нитей, но и у протофибрилл и даже фибрилл. При сильной механической нагрузке такие фибриллы могут раскручиваться, чем достигается некоторое удлинение волокна. Кроме того, спиральное строение увеличивает гибкость волокна, что легко понять, сопоставив гибкость сплетенного из тонких проволок троса с гибкостью сплошного металлического стержня такого же диаметра.
Итак, коллагеновое волокно представляет собой пучок фибрилл, в свою очередь состоящих из протофибрилл, образованных макромолекулами коллагена, то есть тройными полипептидными цепочками.
Следует помнить, что даже рядом лежащие коллагеновые волокна могут иметь различный возраст и отличаться друг от друга по свойствам. В частности, наряду со зрелыми дифференцированными волокнами встречаются более молодые аргирофильные волокна.
Эластические волокна. Если коллагеновые волокна при рассматривании их в световой микроскоп кажутся продольно исчерченными (поперечная исчерченность при этих увеличениях не видна), что отражает их фибриллярное строение, то эластические волокна представляются гомогенными.

Рисунок 18. Схема строения эластического воокна (по Холлу): 1 – белковые молекулы; 2 – полисахарид; 3 - наружный слой.
Они всегда анастомозируют друг с другом, образуя единую эластическую сеть, легко растяжимую и непрочную на разрыв. Толщина волокон крайне разнообразна {от еле различимой под микроскопом до 12μ.). Окрашиваются они только специальными красителями (орсеином и некоторыми другими); клея при варении не дают. Двойное лучепреломление обнаруживают только при сильном растяжении, то есть когда составляющие их частицы вытягиваются в длину (как может вытягиваться мех гармоники) и приобретают упорядоченное линейное расположение. Они очень стойки к кислотам и щелочам и долго сохраняются, например при туберкулезе, в совершенно разрушенной ткани.

Рисунок 19. Электронная микрограмма коллагеновых волокон коровы (по Гроссу и Шмиту). Чередование светлых и темных полос с периодом 640 Å.
Электронная микроскопия показывает, что эластические волокна тоже состоят из нитей белка (проэластина), но их удается видеть только после растворения цементирующего, богатого углеводами вещества (эластомуцина). Из рисунка 19 видно, что в эластическом волокне есть срединная осевая нить из белковых молекул и наружный слой, также представленный белковыми молекулами, но соединенными между собой полисахаридом. Если соответствующими энзимами разрушить этот полисахарид, все волокно распадается.
Значительной сложности эластические элементы достигли в стенке крупных артерий, где они имеют над относительно толстых мембран с коллагеноподобной сердцевиной. С поверхности мембраны одеты мукополисахаридной муфтой, отличающейся особенно активным обменом веществ. Не случайно в области эластических мембран и возникают в первую очередь различные нарушения обменных процессов — отложение липоидов и минерализация.
По мере старения животного химический состав эластических волокон меняется, усиливается их сродство с минеральными солями (обызвествление стенки артерий), волокна становятся хрупкими и легко распадаются на отдельные фрагменты.
Дата добавления: 2015-02-10; просмотров: 572; Мы поможем в написании вашей работы!; Нарушение авторских прав |