КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Случайные процессы при описании популяций
Рассматриваемые выше модели – детерминистские. Это должно иметь какие-то основания, которые мы и попытаемся сейчас обсудить.
Если речь идет о динамике популяций, то можно выделить по крайней мере два аспекта, по которым детерминистская модель не может служить точным отражением реальной экологической системы: во-первых, она допускает бесконечно большую численность популяции; во-вторых, не учитывает случайных колебаний, происходящих в среде во времени.
В качестве примера детерминистской экологической модели рассмотрим уравнение
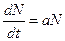 , (10.1)
, (10.1)
где N – число особей в момент времени t,
а – истинная скорость роста.
Решением этого уравнения, удовлетворяющим начальному условию
N(0)=No, (10.2)
является функция
N(t)=N0eat, (10.3)
(так называемый закон Мальтуса – закон роста популяции без конкуренции). В основе главного допущения здесь лежит то, что за короткий промежуток времени t каждая особь порождает aΔt новых особей.
В соответствующей стохастической модели принимается более правдоподобное допущение, согласно которому за период Δt одна особь с вероятностью λ производит одного потомка и с вероятностью μΔt умирает. Обозначим через рi(t) вероятность того, что в момент времени t численность популяции равна i, i = 0, 1, 2, ... Рассмотрим величину pi(t + Δt). В силу малости Δt можно считать, что численность популяции останется прежней, равной i, в результате трех независимых событий – появления потомков в популяции с численностью i–1, отсутствия случаев рождения и смерти в популяции с численностью i и смерти в популяции с численностью i+1. При этом вероятность pi(t + Δt) равна сумме вероятностей этих событий:
pi(t + Δt) = (i-1) λ pi-1 (t) Δt+(1-i(λ+μ)pi(t) Δt+(i+1) μi+1(t) Δt , откуда
 (i-1) λ pi-1 (t)- i(λ+μ)pi(t)+ (i+1) μi+1(t).
(i-1) λ pi-1 (t)- i(λ+μ)pi(t)+ (i+1) μi+1(t).
Переходя в полученном соотношении к пределу при t → ∞, получим систему уравнений Колмогорова
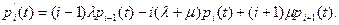 (10.4)
(10.4)
В виде (10.4) уравнения справедливы при i= 2, 3, 4, .... При i = 1 из (10.4) получаем уравнение
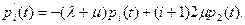 (10.5)
(10.5)
а при i = 0 – уравнение
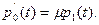 (10.6)
(10.6)
(естественно считать, p-1(t)≡0).
Если в начальный момент времени t=0 в популяции имелось N0 особей, то начальные условия для системы обыкновенных дифференциальных уравнений с постоянными коэффициентами (10.4)–(10.6) имеют вид:
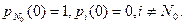 (10.7)
(10.7)
Рассматриваемый процесс гибели и рождения является случайным процессом (классическим примером цепей Маркова [17]), а само решение задачи (10.4)–(10.7) можно получить стандартными методами теории обыкновенных дифференциальных уравнений (см., например, [47]). Нас интересуют следующие вероятностные характеристики: ожидаемое значение, т. е. среднее значение популяции в момент времени t
N(t)= 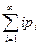 (t) (10.8)
(t) (10.8)
и вариация (дисперсия), т. е. среднее квадратичное отклонение от N(t)
 . (10.9)
. (10.9)
Для вычисления N(t) заметим, что из уравнения (10.5) и первого уравнения из (10.4) вытекает
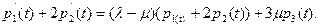
Продолжая этот процесс сложения, получим
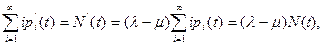
т. е. обыкновенное дифференциальное уравнение
N'(t)=(λ - μ)N(t) (10.10)
с начальным условием (10.7)
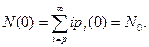 (10.11)
(10.11)
Решение его, очевидно, равно
N(t)= 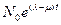 , (10.12)
, (10.12)
в частности, при λ > μ численность популяции экспоненциально возрастет (при λ=μ+a определяется уравнением (10.3)), а при λ < μ экспоненциально убывает при t → ∞. Аналогично (см. [17]) вычисляется вариация
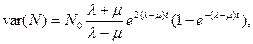 (10.13)
(10.13)
откуда при λ > μ для коэффициента вариации получаем выражение
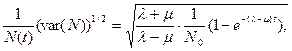 (10.14)
(10.14)
которое при t → ∞ стремится к величине 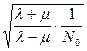 . Следовательно, при достаточно больших начальных значениях популяции N0 среднее квадратичное отклонение от N(t) является равномерно малым, и детерминистская модель дает адекватное представление о поведении популяции при больших значениях времени.
. Следовательно, при достаточно больших начальных значениях популяции N0 среднее квадратичное отклонение от N(t) является равномерно малым, и детерминистская модель дает адекватное представление о поведении популяции при больших значениях времени.
Дата добавления: 2015-01-05; просмотров: 227; Мы поможем в написании вашей работы!; Нарушение авторских прав |