КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Адаптивных сочетаний генов
В настоящее время уже известно много способов видообразо-
вания. Некоторые из них отличаются друг от друга лишь в деталях,
другие различаются принципиально. Дать надежную классифика-
цию способов видообразования пока не удается, поскольку не най-
ден единый принцип. Мы остановимся кратко на двух разных
принципах и приведем некоторые примеры по каждому из них.
I. По характеру изоляции:
1. Аллопатрическое видообразование(allos – разный, patria –
родина). Данный способ базируется на географической изоляции,
разделяющей популяции в пространстве. Отметим, что с эволю-
ционной точки зрения наиболее интересным объектом являются
популяции, находящиеся на периферии видовых ареалов. Дело в
том, что периферия – это граница пригодных для вида условий.
Приграничные жители больше всех прочих подвергаются экстре-
мальным воздействиям внешних факторов и одновременно имеют
наибольшие шансы освоить новые условия с помощью новых
признаков. Вероятно, многие новые виды обособлялись в краевых
популяциях материнских видов.
а) несколько миллионов лет назад Панамский перешеек меж-
ду двумя Америками находился под водой, и на его месте в океане
обитало много видов рыб. Поднявшись, перешеек разделил Ат-
лантический и Тихий океаны. Представители практически каждо-
го тамошнего вида рыб оказались разделены, а условия их жизни
не совпадали полностью. В дальнейшем группы эволюционирова-
ли независимо друг от друга. Сейчас по обе стороны перешейка
обитает 1200 видов рыб. Из этого списка одинаковыми являются
лишь около 70 видов, то есть всего 6%;
б) на Гавайские острова были заселены африканские сухопут-
ные улитки семейства Achatinellidae. Они малоподвижны, и в до-
линах между горными хребтами исходный вид образовал много
непохожих друг на друга изолятов. Сейчас разные специалисты
выделяют на Гавайях от нескольких десятков до 300 новых видов
улиток. На одном лишь острове Оаху в каждой их двадцати пяти
долин свой вид улиток;
в) в разных частях острова Новая Гвинея пять оседлых видов
райских сорок образовались от одного исходного;
г) в ледниковый период большие ареалы многих видов были
разделены ледниками на то или иное количество изолятов, в каж-
дом из которых эволюция шла особым путем и давала отчетливые
различия между группами. С уходом ледников целостность едино-
го когда-то ареала может и не восстановиться. Например, таким
способом образовались пять рас ландыша – европейская, кавказ-
ская, дальневосточная, сахалино-японская и североамериканская.
Три расы – европейская, кавказская и сибирская – сформировались
у купыря лесного ( Anthriscus silvestris). Ареал рыбы вьюн сейчас
представляет собой два изолята в Европе и на Дальнем Востоке;
д) в районе Берингова пролива сформировался некий вид ча-
ек, который расселялся от этого центра по побережью Ледовитого
океана. Расселение шло в двух направлениях – евразийском и се-
вероамериканском. По ходу расселения на разных участках побе-
режья образовалось более двадцати подвидов. Все соседние под-
виды скрещиваются друг с другом. Обе линии расселения
встретились в Северо-Западной Европе, и оказалось, что «окруж-
ность» не может замкнуться, ибо встретившиеся группы не скре-
щиваются. Мы знаем их как серебристую чайку ( Larus argentatus)
и клушу-хохотунью ( Larus fuscus);
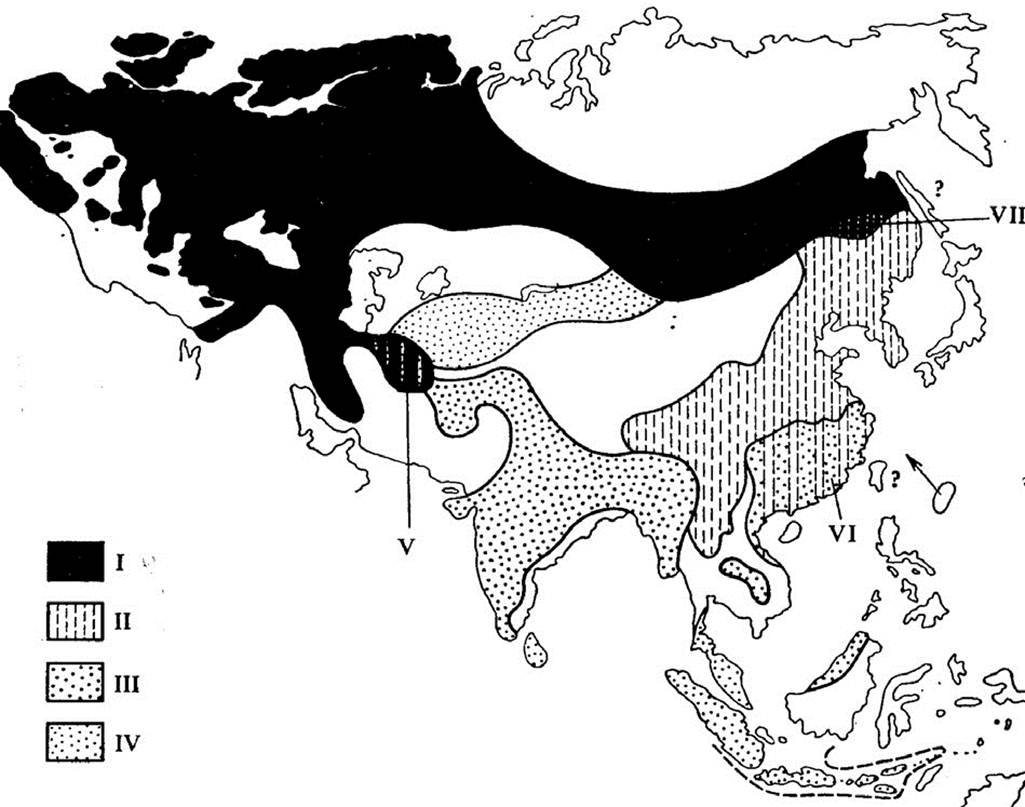
е) аналогично предыдущему случаю, большая синица живет
от Ирландии до Индонезии. В ходе эволюции она расселялась из
Европы на восток по двум направлениям. Северное направление
шло по лесам умеренной зоны, а южное – через Иран, по тропиче-
ским лесам Южной Азии. В обоих направлениях образовывались
подвиды общим числом более тридцати. В бассейне Амура круг
замкнулся, и опять-таки «концевые» подвиды ( Parus major major
и Parus major minor) скрещиваться не способны. Два последних
случая называют кругом рас, либо кольцевым видом (рис. 7).
Рис. 7. Кольцевой ареал большой синицы (Parus major):
I – Parus major major; II – P.m. minor; III – P.m. cinereus;
IV – P.m. boxharensis; V – переходные популяции (major x cinereus);
VI – переходные популяции (cinereus x minor);
VII – перекрывание ареалов major и minor
ж) широко известный пример – население Австралии, Мадага-
скара и Новозеландского архипелага. Длительная изоляция этих
территорий привела к появлению огромного числа эндемичных ви-
дов, отсутствующих в других местах (см. методы изучения эволю-
ции).
Еще раз вспомним, что репродуктивная изоляция как этап
формирования нового вида может возникать и без территориаль-
но-механической изоляции, то есть на одной территории. Отсюда
второй пункт обсуждаемой классификации.
2. Симпатрическое видообразование (sim – вместе, patria –
родина).
а) в рассказе о репродуктивной изоляции мы уже привели це-
лый ряд примеров этого пути видообразования: нескрещиваю-
щиеся нерестовые расы рыб и миног, близкие виды дрозофил и
сосен с разными сроками размножения, виды пауков, отличаю-
щихся лишь брачным танцем, и виды соловьев с разной песней,
светляки с разным характером свечения;
б) у самоопыляющихся растений ястребинок близкие виды
отличаются предпочтением тех или иных субстратов (пески, ска-
листые склоны, засоленные участки почвы);
в) сходным образом олигохеты Байкала дали много симпат-
рических видов со специфической привязкой к микробиотопам
(пескам, илам, камням, обрастаниям), каждый из которых дает
разные типы укрытий и обусловливает формирование характер-
ных морфологических черт;
г) обнаруживается все больше примеров видов-двойников у
насекомых, обитающих на одних территориях. Различия между
видами выявляются в строении копулятивных аппаратов.
По современному состоянию флоры и фауны не всегда просто
определить способ видообразования. Виды, получившиеся алло-
патрическим путем, могут при расселении снова оказываться в
одном ареале или в соприкасающихся. И если не знать истории их
возникновения, можно ошибиться в отношении способов видооб-
разования.
Есть красивый пример с австралийскими мухоловками. У ис-
ходного вида был широкий ареал, вытянутый с запада на восток
континента. При иссушении климата он разбился на два участка, в
одном из которых сформировался новый вид. При изменении
климата в обратную сторону произошло расселение исходной
формы, и ареал нового вида оказался в пределах ареала материн-
ского. Позднее ареал вновь раскололся на два. Теперь на востоке
Австралии мы имеем вторичную симпатрию при аллопатрическом
видообразовании.
Точно так же пресноводная лососевая рыба ленок имеет две
формы – острорылую (бронзовая с черными пятнами) и тупоры-
лую (серебристая с радужными пятнами). В ледниковый период
тупорылый ленок образовался в Джунгарии, а острорылый – на
Дальнем Востоке. В послеледниковое время они расселялись на-
встречу друг другу, и ныне их ареалы перекрываются, давая сим-
патрию при аллопатрическом видообразовании. Подобные ситуа-
ции мы находим при перекрывании ареалов западного и
восточного соловьев в Центральной Европе, а также при много-
кратном вселении на острова, в водоемы и пещеры.
При незавершенном аллопатрическом видообразовании ди-
вергенция видов обратима, и возможны реверсии на разных ее
этапах. Например, в местах соприкосновения ареалов серой и
черной ворон есть узкие зоны гибридизации. В ледниковое время
обособились подвиды снегиря. Один из них ( Pyrrhula pyrrhula
pyrrhula) обитал на северо-востоке Европы и в Северной Азии, а
второй ( P.p. minor) – в Западной Европе. При послеледниковой
встрече подвидов в Центральной Европе образовался подвид
P.p. germanica, являющийся результатом гибридизации. В Италии
есть гибридная популяция воробьев, которая скрещивается на се-
вере с домовым воробьем ( Passer domesticus), а на юге – с испан-
ским ( P. hispaniolensis).
3. Филетическое видообразование. Это особый случай, при
котором популяция постепенно изменяется в определенном на-
правлении без раскола на формы (своего рода эволюция «строго
по прямой»). Данный способ устанавливается лишь палеонтоло-
гическими методами, дающими последовательность ископаемых
форм (см. палеонтологические методы эволюции). Непрерывный
временн ой ряд состояний популяции называют филетической
линией. Выделять филетические линии трудно, поскольку всегда
существует опасность упустить какие-то эволюционные ответвле-
ния. Примеры, приведенные в разделе о методах эволюционной
теории, можно рассматривать лишь для ограниченного отрезка
времени. Рано или поздно происходит раскол на разные виды и на
прямой эволюционной ветви появляются боковые «побеги». Кро-
ме того, при медленном непрерывном изменении состояния груп-
пы почти невозможно установить границы вида. Они всегда будут
условны, ибо трудно сказать, где на филетической линии кончает-
ся один вид и начинается другой.
II. По скорости формирования полной генетической изо-
Дата добавления: 2015-04-16; просмотров: 354; Мы поможем в написании вашей работы!; Нарушение авторских прав |