КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
ПРАКТИКА: КОРЕНЬ. 2 страница
В зависимости от расположения чешуи различают луковицы пленчатые, черепитчатые и сложные (сборные). Пленчатыми луковицами называют такие, у которых внутренние чешуи мясистые, а верхние сухие, широкие и плотно охватывают одна другую (лук, гиацинт). У черепитчатых луковиц наружные чешуи узкие, не налегают одна на другую (лилия). Сложной луковицей называется такая, которая состоит из большого количества деток, луковичек или зубков и сверху одета общими чешуями (чеснок).
На верхушке донца находится верхушечная почка, которая развиваясь, на 2-й год жизни дает надземные стебли. Луковица служит растению для вегетативного размножения. В ней откладываются запасные питательные вещества (сахара, крахмал и др.). Растения, имеющие луковицы, называются луковичными (лук, чеснок, лилия, гиацинт, тюльпан и др.).
Некоторые растения имеют надземные воздушные луковицы. Такие надземные маленькие луковички развиваются у чеснока в соцветиях, а у лилий — в пазухах листьев и служат также для вегетативного размножения.
Корневище. Это подземный побег, который образуется у многолетних растений. По форме и строению существует большое разнообразие корневищ. Внешне корневище часто сходно с корнями, но отличается от них как по морфологическому, так и по анатомическому строению. Корневища растут, как стебли, своей верхушкой. У корневищ часто хорошо заметны узлы, на которых вместо настоящих листьев образуются довольно разнообразной формы чешуи — видоизмененные редуцированные листья. В пазухах чешуи формируются почки, дающие начало надземным и подземным побегам, а из узлов образуются придаточные корни. Верхушка корневища заканчивается почкой, а не чехликом, как у корня. Почки корневищ ежегодно дают новые подземные и надземные побеги. Корневища отличаются очень быстрым ростом, и у некоторых растений (пырей ползучий, осока) они достигают больших размеров (50...300 см). Корневища служат растениям для вегетативного размножения, причем они хорошо развиваются даже из небольших кусочков. Поэтому такие злостные сорные растения, как пырей ползучий, быстро размножаются разрезанными при вспашке корневищами.
У многих растений корневища бывают толстые, мясистые. Они служат местом отложения питательных, дубильных (дубящих), лекарственных, ядовитых веществ (раковые шейки, валериана, вех ядовитый) или имеют вид длинных тонких тяжей (пырей ползучий).
Колючки. Это твердые остроконечные образования, развивающиеся из пазушных почек и представляющие собой надземные видоизменения побега. Колючки могут быть простые и ветвистые, без листьев, или они несут небольшое количество недоразвитых листьев. Такое видоизменение побега является для растений хорошей защитой от животных. В отличие от шипов колючки сдираются только с частью древесины. Они свойственны таким
растениям, как дикая груша, дикая яблоня, боярышник, терновник.
Кладодии, или ф и л л о к л а д и и. Эти видоизмененные надземные побеги внешне напоминают листья (плоские, тонкие), поэтому их называют листовидными побегами. Такие побеги несут на себе редуцированные листья и цветки, чем они и отличаются от настоящих листьев, которые никогда не несут цветков.
Примером такого видоизменения побега могут служить кладодии иглицы (рис.).
Усики. К надземным видоизменениям побега относятся также стеблевые усики. В отличие от усиков листового происхождения они, подобно колючкам и кладодиям, выходят из пазухи листа. Такие усики характерны для представителей семейства тыквенные (огурец, тыква), для винограда; ими растения прикрепляются к опоре.

Рис. Видоизменения надземных побегов:
а — колючки боярышника; б — кладодии иглицы; в — сочные стебли кактуса; / — кладодий; 2 — цветок; 3 — редуцированный лист
С очные стебли. Некоторые растения (кактусы и молочайные) имеют сочные, мясистые стебли, приспособленные для накопления и сохранения большого количества влаги; они обладают специальной водоносной тканью. Листья у таких растений часто превращаются в острые мелкие колючки, а
функции листьев выполняют стебли. Произрастая в засушливых районах, растения с сочными стеблями и листьями очень экономно расходуют воду, так как имеют мало устьиц.
Анатомическое строение стебля. Строение стебля тесно связано с выполняемыми им физиологическими функциями. В стебле, как и в корне, наблюдается первичное и вторичное строение.
Первичное строение стебля. Развитие стебля осуществляется за счет клеток первичной меристему, сосредоточенных в конусе нарастания и образующих 2 обычно хорошо выраженные зоны тунику и корпус.
В стебле первичного строения различают эпидермис, первичную кору и центральный цилиндр. Эпидермис развивается из туники, а если туника состоит из нескольких рядов клеток, она образует и наружные слои первичной коры. Корпус дает начало первичной коре и всей проводящей системе стебля, т. е. центральному, или осевому, цилиндру (стеле).
Эпидермис стебля состоит из вытянутых клеток с утолщенными внешними стенками, часто покрыт кутикулой и имеет небольшое количество устьиц. На поверхности эпидермиса образуются опушение из разнообразных волосков, шипы или восковой налет.
Первичная кора состоит из основной паренхимной ткани. В клетках ее наружных слоев часто содержатся хлоропласты. У многих растений в состав первичной коры входит также механическая ткань — колленхима. Склеренхима в первичной коре встречается очень редко. Внутренние клетки первичной коры образуют кольцо эндодермы, которая у многих растений превращается в крахмалоносное влагалище. В его клетках содержатся многочисленные мелкие крахмальные зерна, которые не потребляются растением даже при значительном недостатке питательных веществ. Некоторые авторы считают, что крахмалоносное влагалище обусловливает вертикальное положение стебля. Клетки эндодермы могут утолщаться и древеснеть, но при этом сохраняются тонкостенные пропускные клетки. У многих растений эндодерма бывает недостаточно хорошо выражена.
Центральный цилиндр состоит из перицикла, граничащего с эндодермой, системы проводящих элементов и сердцевины. Перицикл является боковой первичной меристемой, он может давать начало клеткам камбия, в нем зарождаются придаточные корни и придаточные почки. У многих растений перицикл полностью дифференцируется в механическую ткань — склеренхиму (перициклические волокна) или в клетки основной ткани.
Проводящие элементы центрального цилиндра развиваются из особых участков конуса нарастания — прокамбия. Прокамбий возникает в результате деления некоторых клеток первичной меристемы преимущественно в продольном направлении, тогда как образование между ними поперечных перегородок замедляется. Появляются группы своеобразных узких вытянутых клеток, заполненных густым зернистым содержимым, которые к центру стебля образуют элементы первичной ксилемы, а к его периферии — элементы первичной флоэмы.
Внутренняя часть центрального цилиндра состоит из клеток паренхимной ткани, образующей сердцевину. Наличие сердцевины является характерным признаком стебля, который отличает его от корня. Сердцевина соединена с первичной корой первичными сердцевинными лучами, также состоящими из паренхимных клеток. Сердцевина может полностью или частично разрушаться, и тогда в центре стебля образуется полость.
Прокамбий может закладываться в конусе нарастания по-разному. Способ его заложения и последующее развитие, строение и
|
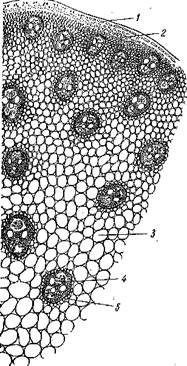
Рис. Строение стебля однодольного растения:
/ — эпидермис; 2 — склеренхима; 3 — основная паренхима; 4 — закрытые проводящие пучки; 5- склеренхимное кольцо

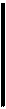 расположение проводящих пучков определяют анатомические особенности стебля у однодольных и двудольных растений.
расположение проводящих пучков определяют анатомические особенности стебля у однодольных и двудольных растений.
Однодольные растения. Особенностью стебля большинства однодольных растений (кукуруза, сорго, купена) является слабое развитие первичной коры, которая обычно состоит из 1...3 рядов клеток, содержащих хлоропласты. Иногда она полностью отсутствует, и склеренхима перицикла (перициклические волокна) непосредственно примыкает к эпидермису в виде непрерывного механического кольца.
Прокамбий в конусе нарастания всегда закладывается в виде отдельных участков (тяжей) и образует изолированные проводящие пучки. Поэтому, характерной чертой всех однодольных растений является пучковое строение стебля. Проводящие пучки, проходя по стеблю, постепенно отклоняются к его центру, а затем снова направляются к периферии. Это обусловливает типичное для однодольных расположение пучков по всей толще центрального цилиндра и отсутствие выраженной сердцевины, вследствие чего на поперечном срезе стебля пучки оказываются разбросанными по всей его поверхности.
В процессе развития проводящих пучков прокамбий полностью дифференцируется в элементы флоэмы и ксилемы и не образует клеток пучкового камбия. Сформировавшиеся проводящие пучки у однодольных всегда закрытые. Стебель растет в толщину только в период деятельности прокамбия и неспособен к дальнейшему утолщению. Наблюдающийся у некоторых древесных однодольных (пальмы, драцены) последующий рост стебля в толщину осуществляется благодаря особому кольцу делящихся клеток, которое образуется из основной ткани.
Значительная часть центрального цилиндра состоит из крупно-клеточной паренхимы, которую пронизывают проводящие пучки, обычно окруженные склеренхимным кольцом.
Стебель большинства злаков (ржи, пшеницы, ячменя и др.) представляет собой соломину и имеет своеобразное анатомическое строение (рис.). На поверхности соломины находится эпидермис, за ним расположена склеренхима, состоящая из толстостенных клеток. Между эпидермисом и склеренхимой находятся участки хлорофиллоносной паренхимы (хлоренхимы), клетки которой имеют тонкие оболочки и содержат хлоропласта. Далее к центру расположена паренхимная ткань, состоящая из крупных тонкостенных клеток, оболочки которых могут древеснеть. Среди клеток паренхимы проходят закрытые коллатеральные пучки. В центре стебля находится крупная полость, окруженная клетками основной ткани.
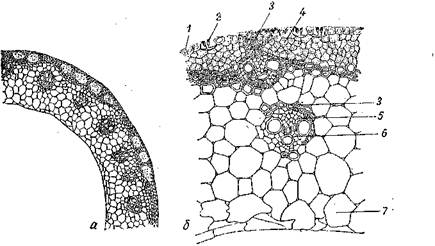
Рис. Строение соломины пшеницы:
а — часть соломины на поперечном срезе при малом увеличении; 6 — то же, при большом увеличении; / — эпидермис; 2 — устьице; 3 — склеренхима; 4 — хлоренхима; 5 — флоэма; 6 — ксилема; 7 — паренхима
Вследствие этого все проводящие пучки как бы сдвинуть к периферии, но на поперечном срезе располагаются рассеянно, как и у других однодольных. Более мелкие пучки сосредоточены у поверхности стебля, более крупные расположены ближе к полости.
Первичное строение сохраняется у однодольных растений в течение всей жизни и наблюдается кик в молодой, так и в более старой части стебля.
Двудольные растения. У двудольных растений разделение стебля на эпидермис, первичную кору и центральный цилиндр выражено достаточно четко, так как первичная кора обычно хорошо развита. Особенностью строения первичной коры является механическая ткань — колленхима, которая в стебле однодольных растений, как правило, отсутствует. Колленхима (уголковая или пластинчатая) чаще залегает непосредственно под эпидермисом, но иногда отделяется от него узкой прослойкой основной ткани. Эндодерма нередко бывает слабо выражена и почти незаметна.
В центральном цилиндре перициклическая склеренхима часто образует сплошное кольцо (тыква) или в виде отдельных участков, разделенных паренхимными клетками, примыкает к флоэме проводящих пучков (подсолнечник, клевер). К центру от склеренхимы расположены первичная флоэма и первичная ксилема, образованные прокамбием.
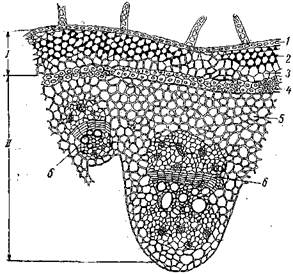
Рис. Первичное (пучковое) строение стебля двудольного растения (тыквы):
/ — первичная кора; // — центральный цилиндр; 1 — эпидермис; 2— уголковая колленхима; 3 — эндодерма; 4 — склеренхима; 5 — основная ткань; 6 — открытый проводящий пучок
Прокамбий у двудольных растений может закладываться как отдельными участками (тыква, клевер, подсолнечник), так и в виде сплошного кольца (лен, табак, древесные растения). В связи с этим первичное строение стебля у двудольных может быть пучковым и непучковым.
Пучковое строение возникает при заложении прокамбия в виде отдельных участков (тяжей), в результате чего образуются, как и у однодольных, изолированные проводящие пучки. Однако в отличие от однодольных эти пучки являются открытыми, так как при их формировании прокамбий дает начало пучковому камбию, за счет которого осуществляется их дальнейший рост. Характерной чертой двудольных растений является также круговое расположение пучков и четко выраженная сердцевина. Это обусловлено тем, что пучки проходят в стебле двудольных всегда на одинаковом расстоянии от его поверхности (рис.). При разрушении сердцевины стебли становятся полыми (тыква, укроп).
Непучковое строение стебля развивается в том случае, если прокамбий залегает в виде сплошного кольца и вместо отдельных пучков сразу образуются кольца флоэмы и ксилемы.
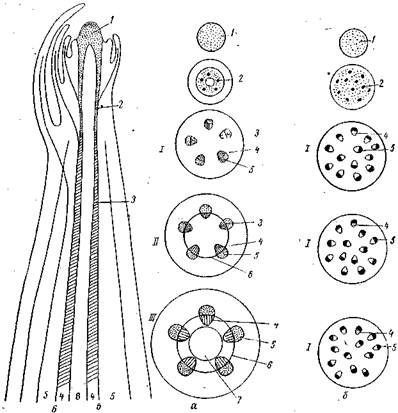
Рис. Схема развития стебля пучкового строения у однодольных и двудольных растений:
а — двудольные; б — однодольные; / — первичное строение; // - формирование вторичного строения; /// — вторичное строение; 1 — первичная меристема конуса нарастания; 2 — прокамбий; 3 — камбий; 4— ксилема; 5 — флоэмa; 6 - межпучковьий камбий; 7 — сердцевина
Первичное строение стебля у двудольных растений наблюдается только в наиболее молодой части стебля — под верхушечной почкой, а далее постепенно заменяется вторичным строением.
Вторичное строение стебля. Развитие вторичного строения тесно связано с особенностями первичного строения стебля и подразделяется на 3 основных тина: пучковый, переходной и непучковый. У травянистых и древесных растений во вторичном строении стебля наблюдаются характерные различия,  обусловленные в первую очередь неодинаковой продолжительностью жизни их стеблей. Травянистый стебель в умеренных широтах является однолетним и ежегодно отмирает в конце вегетационного периода, тогда как древесный стебель — многолетний и может достигать иногда очень большого возраста.
обусловленные в первую очередь неодинаковой продолжительностью жизни их стеблей. Травянистый стебель в умеренных широтах является однолетним и ежегодно отмирает в конце вегетационного периода, тогда как древесный стебель — многолетний и может достигать иногда очень большого возраста.
Строение травянистого стебля. Пучков ы й чип характерен для растений, стебель которых имеет в первичном строении изолированные проводящие пучки (клевер, виноград). При развитии вторичного строения камбий образует элементы вторичной флоэмы и вторичной ксилемы. Клетки основной ткани, разделяющей соседние проводящие пучки, дают начало вторичной меристеме — межпучковому камбию, который дифференцируется в паренхиму сердцевинных лучей. Поэтому, несмотря на образование сплошного камбиального кольца, проводящие пучки и во вторичном строении остаются изолированными. Ввиду того, что у некоторых растений межпучковый камбий бывает слабо выражен, вторичное строение пучкового типа не всегда четко отличается от первичного.
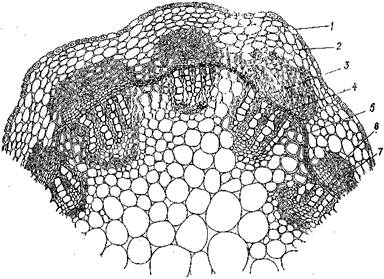
Рис. 78. Пучковый тип строения стебля клевера:
/ — эпидермис; 2 — клетки коры; 3—флоэма; 4— камбий; 5 — межпучковый камбий; 6 — склеренхима; 7 — ксилема
Переходной тип наблюдается у растений, стебель которых в начале развития также характеризуется изолированными пучками, с последующим заложением межпучкового камбия и образованием сплошного камбиального кольца. Отличительная черта переходного типа состоит в том, что вторичную флоэму и вторичную ксилему образует не только пучковый, но и межпучковый камбий, и это приводит к появлению новых (вторичных)* проводящих пучков. Разрастаясь, все пучки постепенно соединяются вместе, и возникают сплошные кольца флоэмы и ксилемы, разделенные камбием (подсолнечник, гречиха, фасоль), т. е. происходит переход от пучкового типа (в первичном строении) к непучковому (во вторичном строении). Поэтому для переходного типа характерно четкое различие между первичным (пучковым) и вторичным (непучковым) строением стебля.
Непучковый тип развивается у растений с кольцевым расположением флоэмы и ксилемы в первичном строении стебля (лен, табак). Вторичная меристема (камбий) также возникает в виде сплошного кольца и образует концентрические слои вторичной флоэмы и вторичной ксилемы. Поэтому при непучковом типе различия между первичным и вторичным строением стебля не выражены.
Строение древесного стебля. Для древесного стебля характерен непучковый тип строения. В связи с тем, что он является многолетним, его отличает от травянистых стеблей способность неопределенно долго расти в толщину, давая прирост каждый вегетационный период. Анатомические особенности заключаются в образовании на его поверхности перидермы, которая сменяет эпидермис, и в появлении четко выраженных годичных колец в древесине.
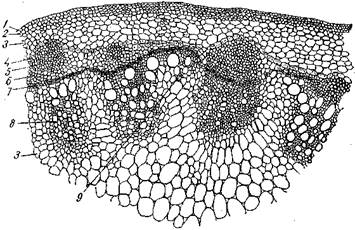
Рис. Переходной тип строения стебля подсолнечника:
/ — эпидермис; 2 — пластинчатая колленхима; 3 — основная паренхима; 4 — эндодерма; 5 — склеренхима; 6 —флоэма; 7— камбий; 8 — ксилема; 9 — пучок, образованный межпучковым камбием
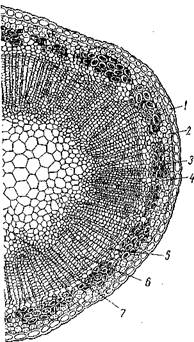
В древесном стебле обычно выделяют кору, камбий, древесину и сердцевину.
В состав коры входят все ткани, расположенные к поверхности от камбия. Наружные слои коры представлены перидермой, состоящей из пробки, пробкового камбия и феллодермы. Иногда на поверхности пробки сохраняются остатки эпидермиса, который через некоторое время, как правило, полностью сбрасывается. За перидермой расположены элементы первичной коры, возникшие в результате дифференциации первичной меристемы конуса нарастания. К ним относятся колленхима и клетки основной ткани, часто содержащие хлоропласты, а также крахмальные зерна и друзы. Ближе к центру стебля находится вторичная кора, образовавшаяся и результате деятельности камбия. Во вторичной коре
|
Рис. Непучковый тип строения травянистого стебля льна:
/ — эпидермис; 2 — первичная кора; 3 — лубяные волокна; 4 – флоэма; 5 – камбий; 6 – ксилема; 7 - сердцевина
четко выражены участки флоэмы, имеющие форму трапеции, в которых лубяные волокна (твердый луб) чередуются с ситовидными трубками, клетками-спутницами и клетками лубяной паренхимы (мягкий луб). Ситовидные трубки в древесном стебле функционируют 2...3 года, затем перестают проводить питательные вещества, отмирают и заменяются новыми ситовидными трубками.
Между участками флоэмы проходят первичные и вторичные сердцевинные лучи, состоящие из клеток основной ткани, в которых часто содержатся питательные вещества — крахмал, жирные масла, сахара. Посредством сердцевинных лучей осуществляется связь между сердцевиной и тканями, лежащими в наружной части стебля.
Камбий состоит из тонкостенных вытянутых клеток прямоугольной формы. Деление клеток камбия в основном осуществляется в тангентальном направлении, т. е. параллельно поверхности стебля, причем элементов ксилемы образуется значительно больше, чем элементов флоэмы. Увеличение числа клеток в камбиальном кольце достигается делением их в радиальном направлении, что обеспечивает неограниченное утолщение стебля. Интенсивность деятельности камбия в течение года неодинакова: наиболее активно он функционирует весной, затем его деятельность постепенно затухает и осенью вообще прекращается — наступает период покоя.
Древесина (ксилема) состоит из сосудов, трахеид, древесинной паренхимы и либриформа. В древесине проходят сердцевинные лучи. В клетках древесинной паренхимы и сердцевинных лучей накапливаются питательные вещества.
Вследствие неравномерной деятельности камбия образованные им клетки древесины имеют различные размеры. Самые крупные клетки образуются весной, когда деятельность камбия наиболее интенсивна. Постепенно деятельность камбия замедляется, и образуемые им клетки становятся более мелкими и толстостенными. Таким образом, за один вегетационный период образуется одно годичное кольцо древесины, в котором хорошо заметны весенние, летние и осенние клетки. После периода зимнего покоя деятельность камбия возобновляется, и формируется новое годичное
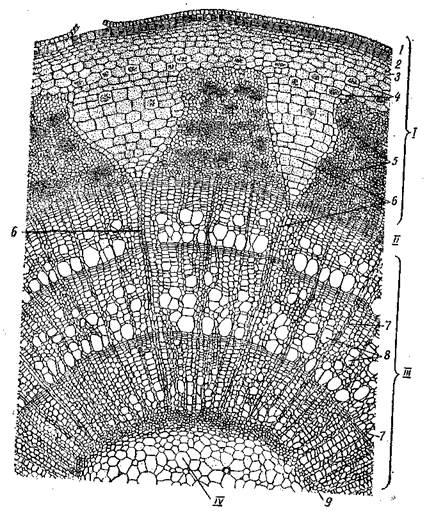
Рис. Непучковый тип строения древесного стебля (разрез трехлетней ветки липы): I – кора; II – камбий; III – древесина; IV – сердцевина; 1 – отмерший эпидермис; 2 – перидерма;3 – колленхима; 4 – клетки основной ткани с друзами; 5 – участки флоэмы; 6 – сердцевинный луч; 7 – осенняя древесина годичного кольца; 8 – весенняя древесина годичного кольца; 9- первичная ксилема
кольцо, крупные весенние клетки которого непосредственно примыкают к мелким клеткам, образовавшимся осенью предыдущего года. Как правило, за год формируется только одно кольцо древесины, однако у некоторых растений (цитрусовые) за один год образуется 2...3 ложных годичных кольца, тогда как стебли растений тропических стран, где нет резкой смены времен года, годичных колец вообще не имеют. Ширина годичных колец зависит от условий произрастания растений.
С возрастом древесина постепенно изменяется и приобретает новые свойства. Сосуды и трахеиды закупориваются тиллами или различными веществами и перестают проводить воду, а клетки основной ткани отмирают. Клетки древесины пропитываются смолами, дубильными веществами, эфирными маслами и нередко интенсивно окрашиваются. Такая отмершая нефункционирующая центральная часть древесины называется ядровой древесиной. Ядровая древесина служит главной опорой ствола, выдерживающей тяжесть кроны. У многих древесных пород (ольхи, тиса и др.) она очень красива и широко применяется как ценный поделочный материал.
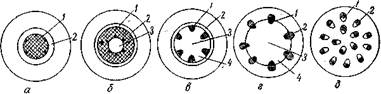
Рис. Схема эволюции стелы:
а — протостела; б — сифоностела; в — диктиостела; г — эустела; д — атактостела; 1 — ксилема; 2 — флоэма; 3 — сердцевина; 4 — прорывы в стеле (сердцевинные лучи)
Периферические слои древесины, расположенные в непосредственной близости к камбию и выполняющие функцию проведения воды с растворенными в ней веществами, называются заболонью. Заболонь обычно имеет более бледную окраску, чем ядровая древесина.
Сердцевина занимает центральную часть стебля и состоит из основной ткани, в клетках которой часто накапливаются разнообразные вещества.
Эволюция стелы. При рассмотрении строения стебля и корня было отмечено наличие характерного для этих органов осевого, или центрального, цилиндра, которым представляет собой систему проводящих, основных и механических тканей, расположенных к центру от первичной коры и отграниченных от нее эндодермой, или крахмалоносным влагалищем. Этот центральный стержень растения получил название стелы, что в переводе означает колонна, столб.
Стела корня мало изменилась в процессе эволюции, тогда как стела стебля прошла в своем развитии ряд этапов. Исходным, наиболее примитивным типом стелы является протостела. Она не имеет сердцевинных лучей и состоит из ксилемы и окружающей ее флоэмы. Эндодерма и перицикл обычно не дифференцированы. Диаметр протостелы очень мал (не превышает 3 мм). Протостела характерна в основном для вымерших растений, из современных растений ее имеют некоторые папоротники.
Несколько более совершенной является сифоностела,
содержащая в центре паренхимную сердцевину и отграниченная
от первичной коры дифференцированней эндодермой. Иногда имеется внутренняя флоэма. Этот тип стелы также характерен для более древних примитивных растений.
Следующим этапом в развитии стелы является д и к т и о стела, для которой характерно возникновение прорывов в массе ксилемы. Эндодерма и перицикл, как правило, хорошо выражены.
Наиболее совершенным типом стелы является э у с т е л а, свойственная голосеменным и большинству двудольных растений. Образуются коллатеральные открытые пучки, отделенные один от другого сильно развитыми сердцевинными лучами. Для однодольных и некоторых двудольных растений характерны разбросанные проводящие пучки на поперечном срезе стебля. Этот тип стелы получил название атактостела.
Во вторичном строении стебель и корень имеют много общего, но в центре стебля всегда находится сердцевина (или полость), а в центре корня расположена первичная ксилема. Переход от первичного строения стебля к первичному строению корня совершается в области подсемядольного колена (гипокотиля) — на участке стебля между семядолями и корневой шейкой. Этот процесс может происходить постепенно, на всем протяжении гипокотиля, что наблюдается у подавляющего большинства растений. Значительно реже он совершается на коротком отрезке гипокотиля. Сущность перехода от строения стебля к строению корня заключается в следующем. Тяжи ксилемы коллатеральных пучков стебля постепенно искривляются, перемещаются относительно продольной оси органа, соединяются попарно и, наконец, сливаются в участки ксилемы. Участки флоэмы вытягиваются в тангентальном направлении и соединяются своими концами, образуя затем компактные тяжи флоэмы, чередующиеся с участками ксилемы.
Особенности строения стебля и продолжительность жизни растений.Строение стебля в значительной степени связано с продолжительностью жизни растений. Существуют растения, которые живут всего 5...7 нед, но есть растения, продолжительность жизни которых исчисляется 5000 лет и более.
Травянистый стебель, как правило, ежегодно отмирает в конце вегетационного периода и весной развивается вновь из почек возобновления, расположенных на зимующих частях растений. Древесный стебель является многолетним и не отмирает в течение всей жизни растения. Деревья, кустарники, полукустарники имеют многолетние древесные стебли. Наибольшей долговечностью отличаются деревья. Ниже приводится примерная долговечность некоторых растений:
Баобаб африканский 5150 лет Кедр сибирский 1200 лет
Мамонтове дерево (секвойя) 5000 » Бук 600... 900 »
Кипарис 3000 » Сосна 500 »
Тис 3000 » Брусника 300 »
Дуб 1200 » Черника 300 »
Ель 1200 » Яблоня 200 »
Растения с травянистым стеблем подразделяют на однолетние, двулетние и многолетние. Однолетние травянистые растения заканчивают свое развитие в течение одного вегетационного периода и к осени полностью отмирают (пшеница, лен и др.). Двулетние растения в 1-й год жизни образуют только вегетативные органы, во 2-й год жизни растения цветут и плодоносят, после чего отмирают (морковь, свекла, брюква, донник и др.). Многолетние травянистые растения имеют долговечные подземные органы (корни, корневища, луковицы), на которых ежегодно закладываются зимующие почки возобновления, развивающиеся весной в надземные побеги (пырей, ландыш, гусиный лук и др.).
Высота растений.Высота стебля у различных растений колеблется в широких пределах — от нескольких сантиметров до десятков метров. Наиболее высокими деревьями являются австралийский эвкалипт, секвойя и дугласова пихта. Эти деревья-гиганты достигают высоты 95...110 м, а отдельные экземпляры— 127 и даже 142 м. Наиболее длинные стебли имеют тропические лианы (до 300м).
Использование стеблей и побегов.Стебли и побеги различных растений широко используются человеком в пищу, в корм животным, как строительный материал, в промышленности для приготовления бумаги, спирта, лекарств и т. д.
ЛЕКЦИЯ 12. Лист.
-
 Лист, функции
Лист, функции - Морфологическое строение. Форма и величина листа. Жилкование. Листья простые и сложные.
- Гетерофиллия (разнолистность)
- Формирование листьев
- Долговечность листьев
- Листопад
- Метаморфозы листьев
- Микроскопическое строение листьев однодольных и двудольных растений. Строение хвоинки.
Общие сведения. Лист представляет собой боковой надземный, а иногда и подземный вырост стебля, вместе с которым образует побег. Через лист у растений осуществляются наиболее сложные процессы по взаимодействию с факторами внешней среды. Наличие листа характерно для высших растений, у которых в результате перехода к наземному образу жизни сформировались под влиянием различных условий окружающей среды большое разнообразие форм листьев и своеобразные их функции.
Лист имеет стеблевое происхождение. Исторически он возник в результате дифференциации и срастания боковых веток стебля в одной плоскости и приспособления к выполнению специфических функций — фотосинтеза, дыхания и транспирации. Поэтому лист, как правило, имеет дорсовентральное строение, т. е. плоскую форму, у которой хорошо различаются спинная — верхняя — и брюшная — нижняя — стороны.
Дата добавления: 2014-12-03; просмотров: 2071; Мы поможем в написании вашей работы!; Нарушение авторских прав |