КАТЕГОРИИ:
АстрономияБиологияГеографияДругие языкиДругоеИнформатикаИсторияКультураЛитератураЛогикаМатематикаМедицинаМеханикаОбразованиеОхрана трудаПедагогикаПолитикаПравоПсихологияРиторикаСоциологияСпортСтроительствоТехнологияФизикаФилософияФинансыХимияЧерчениеЭкологияЭкономикаЭлектроника
Кариотипы древесных растений
Кариотип – хромосомный набор вида, характеризующий их форму, величину и число.
Идеограмма – расположение хромосом в виде диаграммы (Рис.30).
I II III IV V VI VII VIII IX X XI XII
Рис. 30 Кариотип и идеограмма сосны обыкновенной
По характеру кариотипов древесные растения делят на группы:
1. Близкие виды характеризуются разным количеством хромосом и различаются одной, двумя и большим число пар хромосом.
2. Виды некоторых родов представляют собой полиплоидные ряды, когда они отличаются друг от друга кратным числом хромосом.
3. Роды, имеющие виды с однообразным числом хромосом.
4. Роды целого семейства имеют виды с одинаковым числом хромосом.
Все виды семейства сосновых имеют по 12 основных хромосом(n) и по 24 хромосомы в соматических клетках (2n).
У других родов отдельные виды отличаются тем, что содержат хромосомы в несколько раз больше основного числа.
Кариотип различных видов березы образует полиплоидные ряды видов, состоящие из диплоидных (2n=28), триплоидных (3n=42), тетраплоидных (4n=56) и гексаплоидных (6n=84) видов.
У близких родов древесных растений отмечается примерно одинаковое число хромосом: ива и тополь – 19, ольха и береза – 14, бук, дуб, каштан съедобный – 12.
Однако, исходя из числа хромосом, нельзя делать заключение о месте того или иного рода в систематике растений.
Многочисленными исследованиями установлено, что в эволюции растений мутации, связанные с удвоением числа хромосом, имею очень большое значение.
Кариологические исследования позволяют проследить эволюцию видов на фоне меняющихся экологических факторов и географической истории.
Наследственная информация в клетке
Для того чтобы тот или иной компонент мог служить материальной основой наследственности, он должен удовлетворять следующим условиям:
· Выполнять функции, связанные с обменом веществ в клетке.
· Быть способным к самовоспроизведению.
· Точно распределяться между образующимися дочерними клетками.
Таким условиям удовлетворяют хромосомы клетки.
Роль носителя наследственной информации в клетке играетДНК.
Цитологические основы полового размножения.
Мейоз
В процессе развития половые клетки претерпевают мейоз, который состоит из двух последовательных делений:
1. Редукционное деление, уменьшающее число хромосом вдвое (клетки из диплоидных становятся гаплоидными).
2. Эквационное (уравнительное), когда клетки сохраняют гаплоидный набор хромосом (Рис.31).

Рис.31 Последовательные деления мейоза
Цикл мейоза состоит из ряда последовательных фаз, в которых хромосомы претерпевают закономерные изменения
Фазы, относящиеся к первому делению, обозначают римской цифройI,а ко второму –II (Рис.32).
К редукционному делению относят цикл изменений ядра от профазы I до телофазы I, а к эквационному – от профазы II до телофазы II.

Рис. 32 Схема мейоза
Особенностью первого деления является сложная и длительная профаза I которая состоит из ряда последовательных стадий (Рис.33).

Рис. 33 Стадии профазы I
Профаза I самая продолжительная, в которой происходит спирализация хроматина в двухроматидные хромосомы, а центриоли расходятся к полюсам (Рис.34). Затем идет сближение (конъюгация) и укорочение гомологичных хромосом с последующим перекрестом и обменом гомологичными участками (кроссинговер), а также растворение ядерной оболочки.

Рис. 34 Профаза I
В стадии лептонемы сетчатая структура интерфазного ядра переходит в состояние отдельных тонких нитей хромосом (Рис.35).

Рис. 35 Лептонема
Нити хромосом неясно спирализованы и их половинки тесно прилипают друг к другу.
В зигонеме хромосомы начинают притягиваться друг к другу сходными участками (Рис.36).

Рис. 36 Зигонема
Соединение начинается с концов (иногда с центромер). Это взаимное притяжение называют конъюгацией или синопсисом.
На стадии поздней пахинемы и следующей стадии диплонемы происходит перекручивание хромосом, называемое кроссинговером (Рис.37).

Рис. 37 Пахинема
Между гомологичными хромосомами происходит обмен участками.
Диплонема - стадия профазы первого деления мейоза, в течение которой спаренные гомологичные хромосомы расщепляются на хроматиды и образуют тетрады (Рис.38).
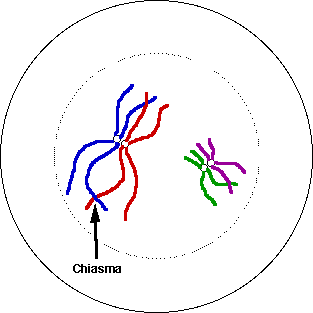
Рис.38 Диплонема
Для последней стадии профазы дикинеза, характерно сильное укорочение и утолщение хромосом за счет их максимальной спирилизации. Биваленты обособляются, и число их в клетке можно сосчитать. Оно оказывается равным гаплоидному (Рис.39).

Рис 39.Дикинез
В этой стадии исчезают ядрышки и ядерная оболочка. Так заканчивается профаза I.
Далее следуют: метафаза I, анафаза I, телофаза I.
Центромеры гомологичных хромосом ориентируются в плоскости экватора веретена деления, что соответствует метафазеI (Рис.40).

Рис. 40 Метафаза I
В анафазеIгомологичные хромосомы бивалентов расходятся к полюсам, вследствие этого число хромосом в дочерних ядрах уменьшается вдвое (Рис.41).

Рис.41 Анафаза I
Следующей фазой первого мейотического деления является очень короткая по продолжительности телофаза (Рис.42).

Рис. 42 Телофаза I
После телофазы I не всегда наступает цитокинез, чаще он наступает после второго мейотического деления. Если цитокинез не наступает после первого деления, то два гаплоидных ядра остаются в одной клетке (Рис.43).

Рис. 43 Цитокинез
Фаза между двумя делениями мейоза называется интеркинезом
В интеркинезе, разделяющим первое и второе деление мейоза, в отличие от интерфазы не происходит репродукции хромосом. Они уже удвоенны и состоят из сестринских хромосом,которые в профазеIIостаютсятакже с двойной природой, то есть состоят из полухроматид. Это указывает на то, что репродукция хромосомных нитей, произошедшая еще в интерфазе, перед началом мейоза, обеспечила хромосомы полухроматидами. Поэтому профаза II не отличается от профазы митоза (Рис.44).

Рис. 44 Профаза II
В метафазеIIхромосомы выстраиваются центромерами в экваториальной плоскости (Рис.45).

Рис. 45 Метафаза II
В анафазеIIосуществляется разделение центромер, и каждая хроматида становится самостоятельной хромосомой (Рис.46).

Рис.46 Анафаза II
ТелофазаII очень сходна с телофазой митоза. Хромосомы деспирализуются, растягиваются и после этого плохо различимы. Нити веретена исчезают, а центриоли реплицируются. Вокруг каждого ядра, которое содержит теперь половинное (гаплоидное) число хромосом исходной родительской клетки, вновь образуется ядерная мембрана. В образование клеточной стенки из одной исходной родительской клетки получается четыре дочерних клетки (Рис.47).

Рис. 47 Телофаза II
В результате первого мейотического деления образуется два ядра с половинным, или гаплоидным числом хромосом. Во втором делении каждое дочернее ядро вновь делится, но в данном случае расходятся хромосомы, образовавшиеся по типу митоза из сестринских хроматид.
Дата добавления: 2014-12-30; просмотров: 2140; Мы поможем в написании вашей работы!; Нарушение авторских прав |